从天文学和生物学角度看
创世论
导言
主张进化论的科学家往往认为创世论缺乏经验支持和科学严谨性。他们认为,创世论不应该被纳入科学课程,因为它无法为地球上生命的多样性和复杂性提供有科学依据的解释。
另一方面,进化论也存在着空白和未解之谜,尤其是在生命起源和生物系统的复杂性方面。自然选择和突变不足以解释在生物体中观察到的复杂结构和功能。此外,进化论只适用于现有的生物体,并不涉及生命的起源。此外,进化论在很大程度上依赖于假设和推测性的重构,从而对其作为生命多样性全面解释的有效性提出了挑战。
本书通过讨论宇宙的创造、地球的独特性以及生命的起源,探讨了创世与进化之间的争论。
在第一部分中,我们将介绍宇宙的等级结构,并讨论天文观测所揭示的宇宙创世过程。然后,我们将研究《圣经》中描述的宇宙创世是否与天文发现一致,地球的年龄是否为6000年,并进一步探讨宇宙的微调性质。
第二部分介绍了有关地球的十个惊人事实,强调了地球支持生命的独特适宜性,并指出了有目的设计的证据。
第三部分探讨生命的起源,挑战传统的进化论,强调生物系统的复杂性是神创造的证据。对“达尔文进化论”一词的适当性进行了研究,随后对人类是否由猿进化而来进行了调查。此外,还介绍了智能设计的概念,并通过讨论粒子物理学、外星生命的存在、动物的本能以及自然界中的数学来探讨创世。
本书最后发出了诚挚的信仰邀请,鼓励读者反思自己的灵性之旅,思考信仰的改变力量。书中介绍了福音,并就如何拥抱信仰提供了实用指导,包括理解和接受永生的步骤,为那些寻求与上帝建立更深联系的人提供了希望和保证。
我希望这本书能让你重新认识创世,加深你对宇宙中错综复杂的设计和目的的理解,并提供一个机会,让你冥想上帝无穷的恩典、智慧和力量,这位神圣的造物主支撑着万物,并邀请我们惊叹于他的杰作。
宇宙的创造
孩提时代,你可能会回忆起在郊外或高山上露营的夜晚,凝视着无数的星星在广袤无垠的天空中闪烁,或惊叹于流星在漆黑的天空中优雅地划过。这样的经历常常让我们充满敬畏和惊叹,深刻体会到宇宙的浩瀚和美丽。在这些时刻,你可能会感受到与宇宙的深刻联系,同时对自己在宇宙中的位置感到谦卑。你的脑海中可能会浮现出一些问题:天空中有多少颗星星?我们的世界之外会有生命吗?宇宙是如何开始的,又是如何结束的?谁创造了这一切?夜空令人惊叹的美丽和神秘的本质激发了人们的好奇心,引人思考宇宙的起源和我们在其中的目的。这些令人着迷的时刻会留下永恒的印记,激励我们去寻找生命中最伟大奥秘的答案。在本章中,我们将从天文学和《圣经》的角度探讨宇宙的起源。我们将通过比较这两种观点,为《创世纪》中的创世记录提供科学支持。此外,我们还将研究地球和太阳哪个先被创造、地球是否有6000年的历史以及微调宇宙的概念。
a.宇宙的等级结构
要讨论宇宙的起源,让我们先来探索宇宙的层次结构。我们将从太阳系开始,依次讨论银河系、外星系、星系团、超星系团和超星系团复合体。
i.太阳系
太阳系由一颗叫做太阳的恒星、围绕太阳运行的八颗行星、火星和木星之间的小行星带、柯伊伯带以及最外层的奥尔特云组成。恒星的定义是以核聚变为动力的自发光天体,而行星则是反射恒星光线的天体。
地球是距离太阳的第三颗行星。从地球到月球的距离是384000公里,乘飞机以每小时1000公里的速度飞行需要16天。地球到太阳的距离约为1.5亿千米,即一个天文单位(AU),乘坐飞机需要17年。到海王星的距离是30AU,柯伊伯带是30到50AU,奥尔特云是2000到200000AU。以光速计算,从地球到太阳需要8.3分钟,到海王星需要4小时,到达奥尔特云内缘需要9.5个月(0.79光年)。如果乘坐飞机,则需要大约85万年。
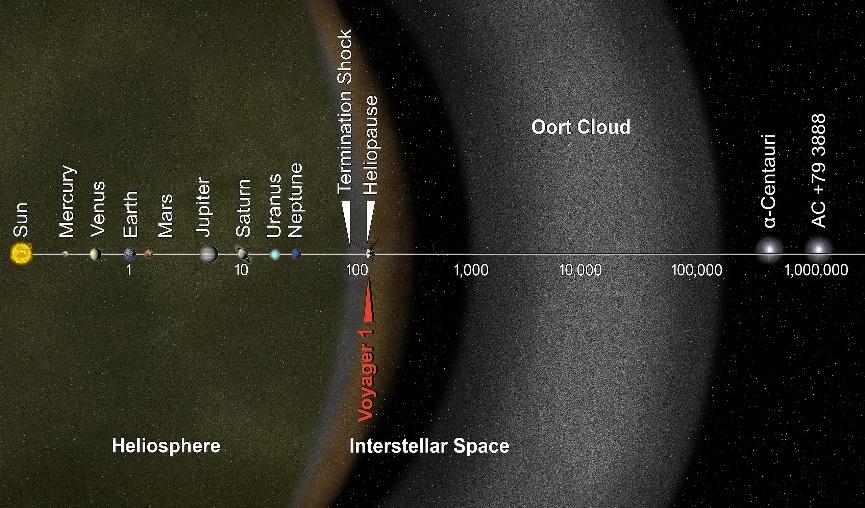
图1.1.包括柯伊伯带和奥尔特云在内的太阳系
彗星可分为短周期彗星和长周期彗星。柯伊伯带是短周期彗星的来源,而奥尔特云则是长周期彗星的来源。由于彗星的起源,它们的轨道高度椭圆,偏心率很大。太阳的大小是地球的109倍,质量是地球的333000倍,自转周期约为25天。
ii.恒星系统
离开奥尔特云,就进入了恒星的世界。距离地球最近的恒星是比邻半人马座,它的大小是太阳的14%,质量是太阳的12%,距离地球约4.2光年。乘飞机前往那里大约需要460万年。
如果你仔细观察夜空中闪烁的星星,就会发现它们有不同的颜色。恒星的颜色取决于它的表面温度:温度较低的恒星呈现红色,而温度较高的恒星则呈现白色。例如,参宿四(αOri)是红色的,太阳是黄色的,而夜空中最亮的恒星天狼星(αCMa)则是蓝白色的。

图1.2.恒星呈现出各种颜色
恒星的质量决定了它的核聚变速度,而核聚变速度又决定了它的光度和寿命。质量较大的恒星比质量较小的恒星消耗燃料的速度更快。恒星以白矮星、中子星或黑洞的形式结束生命。核心质量小于1.4个太阳质量的恒星会变成白矮星,核心质量在1.4到3个太阳质量之间的恒星会变成中子星并以超新星的形式爆炸,核心质量大于3个太阳质量的恒星在经过中子星阶段后会变成黑洞。超新星爆炸的残余物可以循环利用,形成新的恒星。
通常情况下,在理想条件下,城市中肉眼可见的恒星不到100颗,而在乡村中肉眼可见的恒星约有1000颗。这些恒星大多位于距离地球50光年以内的地方。
iii.我们的银河系
银河系是一个棒状螺旋星系,包含2000到4000亿颗恒星以及大量气体、尘埃和暗物质。它的直径约为100000光年,厚度约为1000光年,是一个相对平坦的盘状结构,中央有一个隆起。

图1.3.我们的银河系
太阳距离银河系中心大约2.6万光年,每2.2亿年绕银河系中心运行一圈,这个周期被称为银河年。我们的太阳系位于猎户座支点附近,猎户座支点是位于人马座旋臂和英仙座旋臂之间的一个小旋臂。这个位置距离银河平面约60光年,为从多个方向观测宇宙提供了有利的视角,而银河盘内密集的尘埃和气体对观测的阻碍最小。
iv.星系、星系团和超星系团
仙女座星系(M31)是距离银河系最近的星系,位于,距离地球约250万光年。从北半球用肉眼就能看到它(目视星等=3.4mag),其形状与银河系相似。仙女座星系正以每秒约110公里的速度接近银河系,预计将在大约40亿年后与银河系相撞。
星系大致可分为三大形态类别:螺旋星系、椭圆星系和不规则星系。当两个螺旋星系发生碰撞时,它们之间的引力相互作用会导致巨大的转变,通常会形成一个椭圆星系。这一过程通常会经历多个阶段,包括星系相互作用阶段,然后是红外发光星系(LIRG)或超高亮度红外光星系(ULIRG)阶段。

图1.4.螺旋星系、椭圆星系和不规则星系
如果引力结合的星系少于50个,它们就被称为“星系群”;如果结合的星系有数百或数千个,它们就被称为“星系团”。附近的40多个星系,包括银河系和仙女座,都属于本星系群。本星系群和室女座星系团是室女座超星系团的一部分,而室女座超星系团又是拉尼亚凯亚超星系团的一部分。
超星系团复合体,又称星系丝或超星系团链,是宇宙中一个巨大的大尺度结构,由无数星系超星系团组成,这些星系超星系团由星系、气体和暗物质组成的巨大网络相互连接。这些相互连接的区域形成了网状结构,是宇宙中已知存在的最大结构。它们横跨难以置信的距离,从数亿光年到数十亿光年不等,使较小的宇宙结构相形见绌。其中,海格力斯北冕长城是已知最大的超星系团,它的规模之大令人敬畏。在可观测到的宇宙中,估计有2000亿个星系,分布在约930亿光年的惊人距离上,每个星系都为错综复杂的宇宙结构做出了贡献。
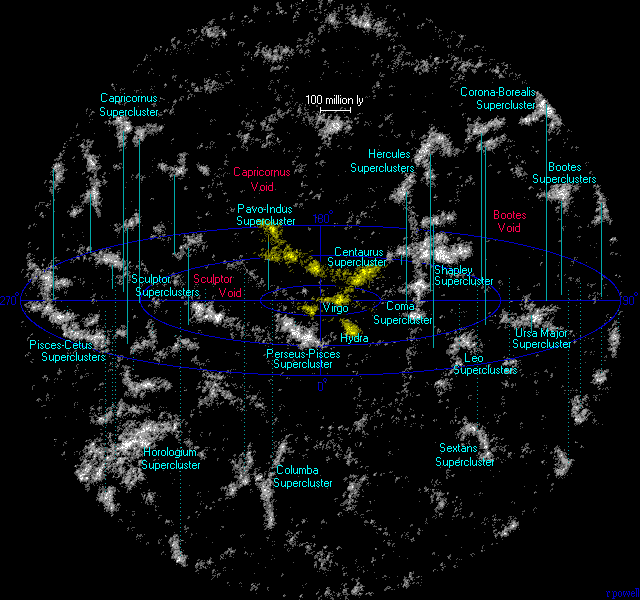
图1.5.附近的超星系团(黄色:Laniakea超星系团)
b.宇宙的创造
宇宙是如何开始的?它一直存在,还是上帝创造的?为了探讨这个问题,我们将研究天文学观测到的宇宙起源以及《圣经-创世纪》中的描述。
i.天文学中的宇宙创生
关于宇宙起源的最广泛支持的理论是大爆炸理论,该理论认为宇宙大约始于138亿年前,当时宇宙是一个非常炙热和致密的点,并迅速膨胀。这自然引出了一个耐人寻味的问题:“在宇宙大爆炸之前?”一种主要的假说认为,在宇宙大爆炸之前,宇宙存在于真空中的量子涨落状态,这是一个动态和概率基础,我们的宇宙正是在这个基础上产生的。
在保罗狄拉克之前,真空被认为是空无一物的空间。1928年,狄拉克结合量子力学和狭义相对论,描述了电子在相对论速度下的行为。有趣的是,该方程提出了电子的两种解法:一种是正能量电子的解法,另一种是负能量电子的解法。狄拉克提出,真空并不是一个空的空间,而是充满了无数个具有负能量的电子(正电子)。因此,真空有时被称为狄拉克海。
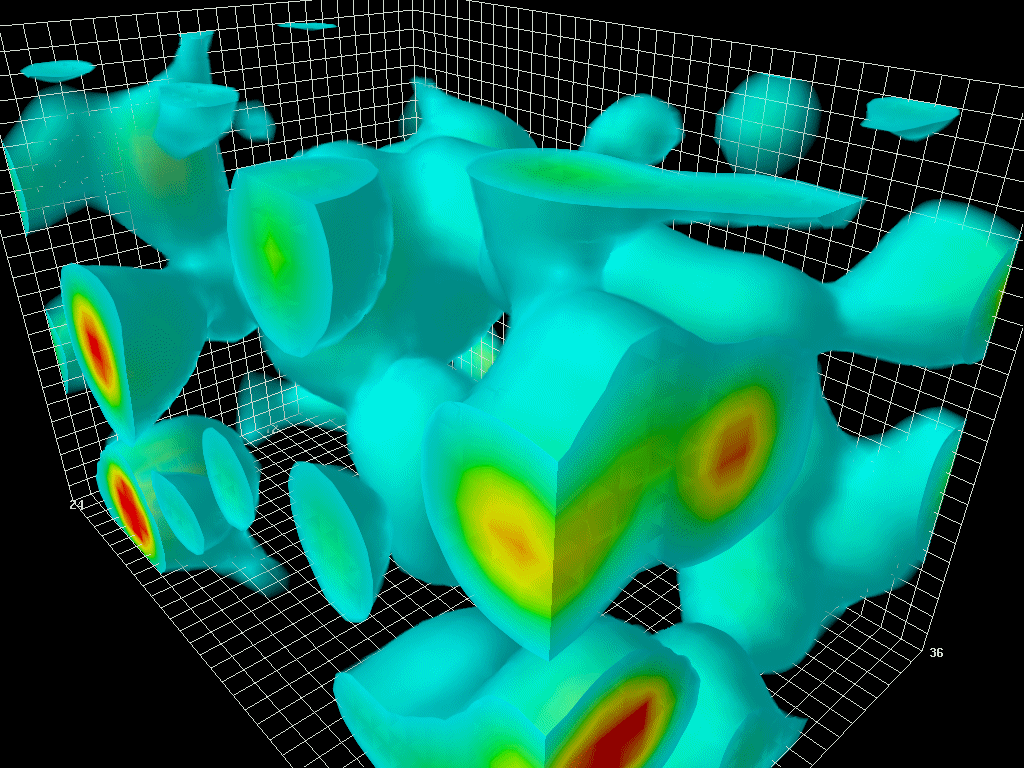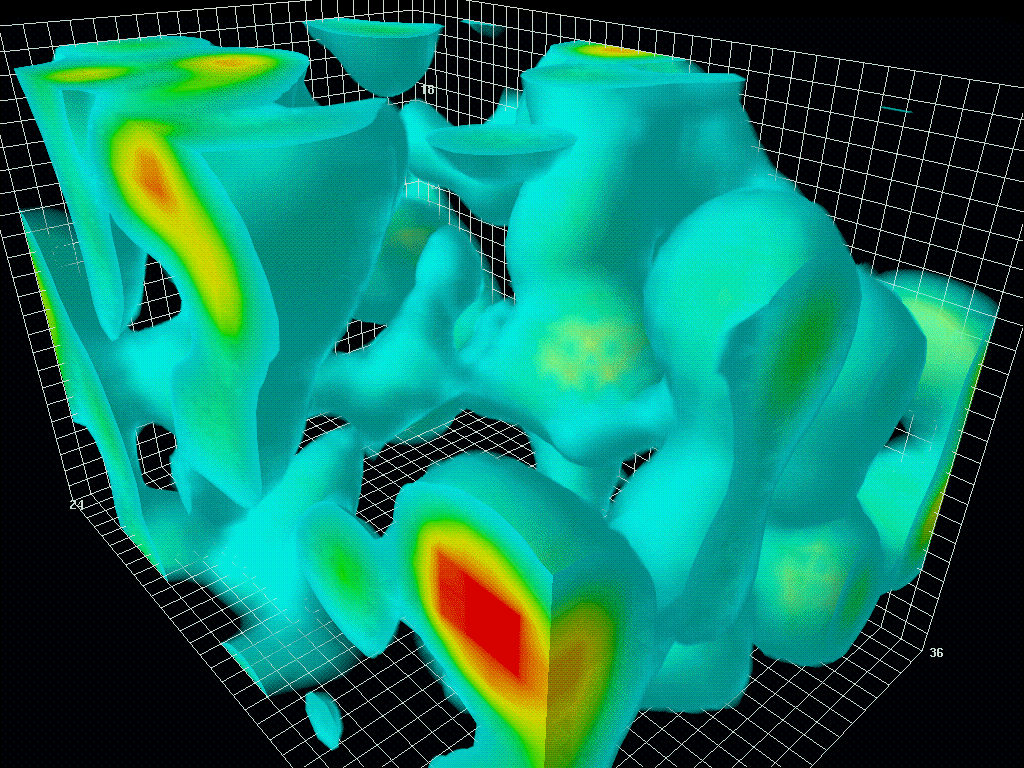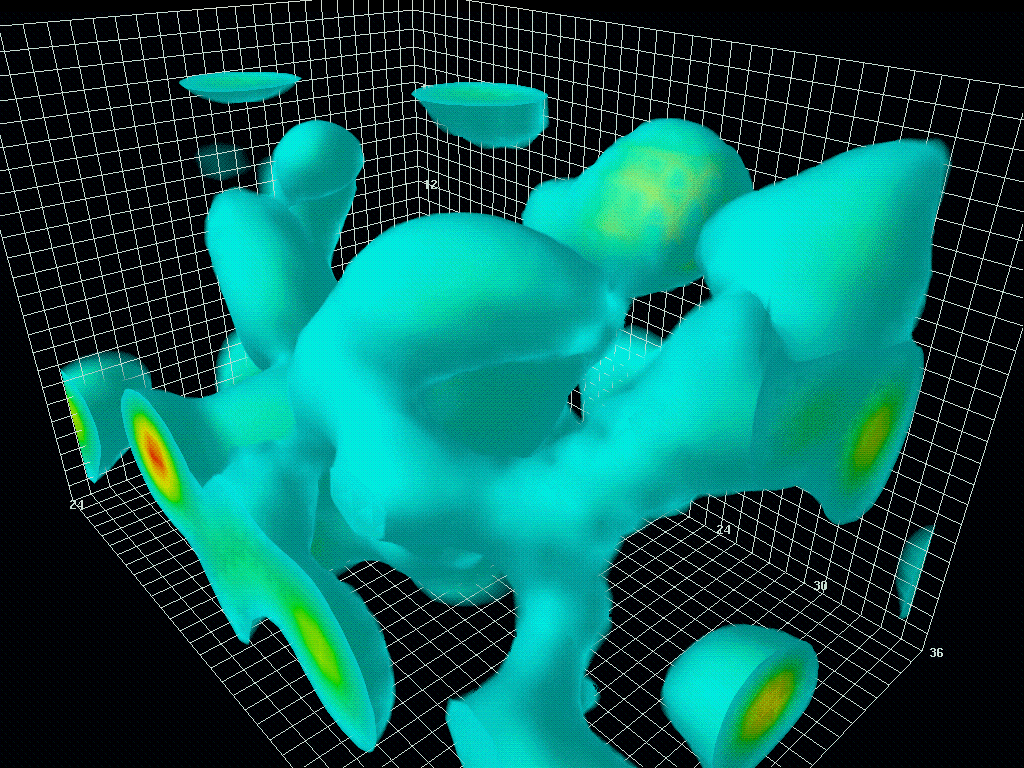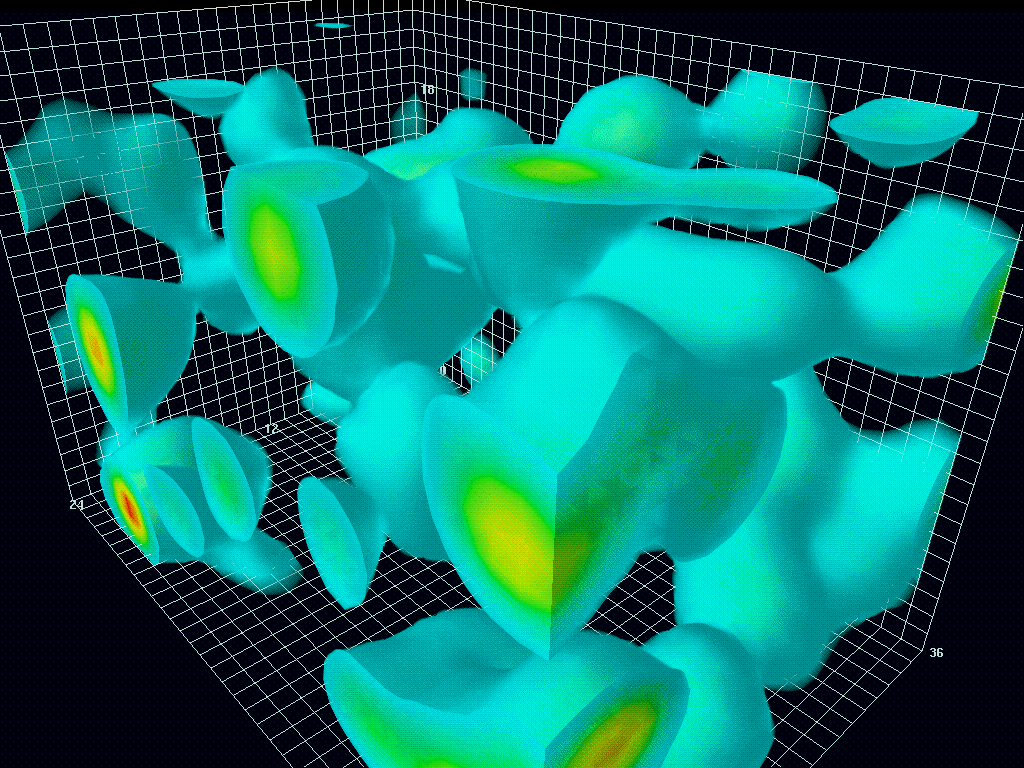
图1.6.真空中量子涨落的三维模型
虽然“狄拉克海”看起来是静态的,但由于海森堡的不确定性原理,它从来都不是静态的。粒子对和反粒子对以随机的方式自发地出现(粒子对产生)和消失(粒子对失效)。时间尺度为10-21秒,人眼无法看到,但如果有相机可以捕捉到,就会像看到波动的大海一样。这就是所谓的量子涨落。宇宙大爆炸就是从量子涨落的海洋中的一个奇点出现的。大爆炸本身就是宇宙的开端。
宇宙大爆炸后,由于温度和密度极高,宇宙立即发生了急剧变化。从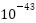 秒(普朗克时间)到
秒(普朗克时间)到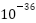 秒,宇宙受大统一理论支配,标准模型中的三种力(强力、弱力、电磁力)被统一。从秒到
秒,宇宙受大统一理论支配,标准模型中的三种力(强力、弱力、电磁力)被统一。从秒到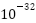 秒,宇宙经历了暴胀纪;从秒到
秒,宇宙经历了暴胀纪;从秒到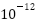 秒,宇宙经历了电弱纪;从秒到
秒,宇宙经历了电弱纪;从秒到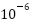 秒,宇宙经历了夸克纪;从秒到1秒,宇宙经历了强子纪;从1秒到10秒,宇宙经历了轻子纪。
秒,宇宙经历了夸克纪;从秒到1秒,宇宙经历了强子纪;从1秒到10秒,宇宙经历了轻子纪。
在轻子纪末,发生了一个戏剧性的关键事件。主要由电子和正电子组成的轻子对和反轻子对发生了相互湮灭。这一过程释放出大量光子(光粒子),使宇宙中充满了光。这些光子成为宇宙中最主要的能量形式,标志着所谓光子纪元的开始。这个时代从宇宙大爆炸后的10秒钟到38万年不等,其特征是自由电子、原子核和光子组成的炙热、致密的等离子体。在此期间,光子被自由电子和质子散射,使它们无法自由传播,从而使宇宙变得不透明。
光子时代结束后,重组时代随之到来,这一时期发生了另一个重要事件。电子与质子结合,形成中性的氢和氦。这是物质主导时代的开始。当这一切发生时,充满等离子体的宇宙逐渐变得透明,变成了我们可以称之为天空的空间。当这种情况发生时,在光子时代产生的、但之前被等离子体限制的光子现在可以在透明的宇宙中自由移动。这些自由移动的光子被观测到时是非常明亮的光,形成了宇宙微波背景辐射。
我们今天看到的恒星和星系就是由重组时期产生的原子形成的。此后,宇宙在大爆炸之后继续膨胀。当宇宙诞生98亿年时,暗能量开始占据主导地位,标志着暗能量主导时代的开始。在这个时代,宇宙继续加速膨胀。这种加速膨胀就是宇宙目前的状态。
ii.宇宙的命运(又是大爆炸?)
宇宙的命运取决于其整体密度。根据WMAP的测量结果,宇宙目前的密度大约等于临界密度(约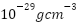 ),误差在0.5%以内。然而,这种不确定性意味着,在获得更精确的测量结果之前,我们还无法确定宇宙的最终命运。如果宇宙的密度大于临界密度,引力最终将战胜宇宙的膨胀,导致宇宙坍缩回自身,这是封闭宇宙特有的灾难性事件,被称为大坍缩(Big Crunch)。
),误差在0.5%以内。然而,这种不确定性意味着,在获得更精确的测量结果之前,我们还无法确定宇宙的最终命运。如果宇宙的密度大于临界密度,引力最终将战胜宇宙的膨胀,导致宇宙坍缩回自身,这是封闭宇宙特有的灾难性事件,被称为大坍缩(Big Crunch)。
相反,如果密度小于临界密度,宇宙就会永远加速膨胀下去,导致一种被称为“大裂解”的情况,这是开放宇宙的特征。在这种情况下,宇宙的温度会随着膨胀的进行而逐渐冷却,恒星的形成最终会因为恒星产生所需的星际介质耗尽而停止。随着时间的推移,宇宙将变得越来越黑暗和寒冷,这个过程通常被称为“热死”。
现有的恒星将耗尽燃料,停止发光。随后,质子衰变会如大统一理论所预测的那样发生,此时宇宙的年龄约为1032年。大约1043年后,黑洞将开始通过霍金辐射蒸发。在所有重子物质衰变和所有黑洞蒸发之后,宇宙将充满辐射。宇宙的温度将冷却到绝对零度,一切都是黑暗和空洞的,类似于宇宙大爆炸前量子涨落的状态。
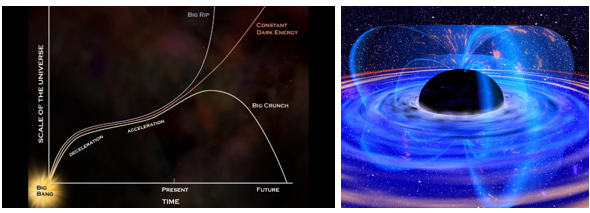
图1.7.宇宙和蒸发黑洞的命运
最近,在距离地球70亿光年的北斗七星方向发现了两个宇宙巨型结构。2022年发现的“巨弧”和2024年发现的“大环”挑战了宇宙学原理,即宇宙在大尺度上是均匀和各向同性的。这些巨型结构需要一个适当的解释。一种可能的解释是,它们是巨大的宇宙弦,或者是上一次大爆炸中超大质量黑洞(霍金点)霍金蒸发的残余物。
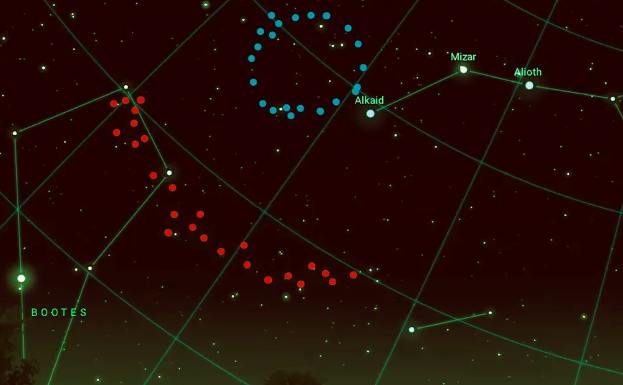
图1.8.大环(蓝色)和大弧(红色)
这种解释与罗杰·彭罗斯的“保形循环宇宙学”(CCC)有关。CCC是一种基于广义相对论的宇宙学模型,在该模型中,宇宙永远膨胀,直到所有物质衰变并留下黑洞。在CCC模型中,宇宙会无限循环往复,在不断膨胀的当前大爆炸中会出现新的大爆炸。

图1.9.共形循环宇宙学
个人而言,我认为CCC具有吸引力,因为它为星系演化中的某些问题提供了潜在解决方案。黑洞质量与恒星速度分散(M-σ关系)之间存在相关性。根据这一关系,黑洞质量约为其所在星系质量的0.1%。最近,钱德拉和JWST通过引力透镜效应发现了一个引人注目的星系UHZ1。UHZ1距离我们132亿光年,当我们的宇宙仅为当前年龄的3%时被观测到。UHZ1的黑洞质量估计值大于其宿主星系的质量。这一超大黑洞质量无法用当前黑洞质量理论解释,但可由CCC理论解释。这可通过以下假设理解:UHZ1中的黑洞是前一次大爆炸中回收的黑洞,并在当前大爆炸中成为UHZ1的种子黑洞。
我们不知道新的大爆炸是如何发生的,而当前的大爆炸仍在膨胀。我们可以尝试使用超空间的概念。在这种情况下,宇宙正在向三维空间膨胀。然而,把我们的三维宇宙想象成一个嵌入高维空间(超空间)的表面。这个高维空间可能是一个四维空间(或更高),我们的整个宇宙只是其中的一个“切片”或“鹤膜”。
随着我们的宇宙不断膨胀,它最终可能会汇聚到这个高维超空间中的一个奇异点,就像二维曲面在三维空间中弯曲并汇聚到一个点一样。超空间中的这个点可能类似于克莱因瓶的瓶颈,是一个表面回环的高维形状。
当宇宙在三维空间中的膨胀汇聚到超空间的这个奇点时,可能会产生能量密度变得极高的条件。如果超空间中的这个奇点无法容纳从当前膨胀的宇宙中涌入的巨大能量和真空能,就会导致爆炸。这次爆炸将是新的大爆炸的开始,从而创造出一个新的宇宙。
这样,不断膨胀的当前大爆炸宇宙可能会在超空间框架内形成一个新的宇宙,而汇聚到一个奇异点则成为CCC循环之间的桥梁。这种高维汇聚为大爆炸的连续循环提供了一种机制,而当前的宇宙仍在膨胀,这种膨胀宇宙的能量也可能为驱动其加速的暗能量做出贡献。
iii.圣经中的宇宙创世
在本节中,我将从天文学的角度探讨《圣经》中描述的宇宙创,研究《圣经》的记载如何与现代科学的理解相一致。这一分析将深入探讨圣经记载与天文观测之间可能存在的相似之处。虽然这种方法提供了一个有趣的视角,但重要的是要认识到还有其他方法来解释《圣经》中的创世记。这些解释可以根据神学、哲学和文化背景而有所不同,每种解释都对宇宙起源的深刻叙述提供了独特的见解。
a)上帝宣布创造宇宙
《圣经》第一卷《创世纪》描述了宇宙的创世过程。
“起初,神创造天地。”(创世纪1:1)
这节经文介绍了上帝的创造行为,断言他是万物的始作俑者。“天地”一词包含了所有的创造,表明了宇宙的整体性。
“地是空虚混沌,渊面黑暗;神的灵运行在水面上。”(创世记1:2)
这里的“地”代表了上帝后来塑造的有形的物质创造物(即重子物质)。“空虚”在英文圣经中被翻译为“void”,意指没有任何事物存在的真空状态。“混沌”则被翻译为“without form”,表示尚未形成任何物质的状态。将两者结合,表明在太初之时,世界处于没有任何事物存在的真空状态。下一句“渊面黑暗”具有深刻的含义。“黑暗”在希伯来语中是חֹשֶׁך(choshek),字面意思是完全的黑暗,没有任何光亮。“渊面”在希伯来语中是תְּהוֹם(tehom),源于הום(hom),意为“骚动”或“波动”。因此,“地是空虚混沌,渊面黑暗” 可以被解释为描述宇宙从一个黑暗且充满波动的真空状态中起源。这种解释与宇宙在其最早阶段——即大爆炸之前——的状态高度一致,当时宇宙以一个处于量子波动中的真空形式存在。
b)光的创造
创世第一天的主要事件是光的创造。
“神说:要有光,就有了光。”(创世纪1:3)
经文指出,上帝通过创造光开启了宇宙的创造。同样,大爆炸始于一系列快速的时代,这些时代总共持续不到一秒钟,最终在光子时代创造了光(光子)。《创世记》1章3节中光的创造与光子时代的光的创造惊人地一致,这有力地将圣经记载与早期宇宙中的这一关键时刻联系起来。
c)创造天空
创世第二天的主要事件是创造天空(天)。
“神就造出空气,...神称空气为天。... 。”(创世记1:7,8)
《创世纪》中描述的创世与大爆炸宇宙学中的重组纪元相关联。在这个纪元之前,宇宙是不透明的,充满了由电子、中子、质子和光子组成的致密、炙热的等离子体。这种等离子体散射光子,阻止它们自由传播,使宇宙对辐射不透明。在此期间,宇宙的直径约为10光年,这意味着没有清晰的空间来形成可见的“天空”。
然而,在重组纪元,宇宙冷却到足以让电子和质子结合,形成中性氢原子。这一过程清除了等离子体,使宇宙变得透明,光子得以在太空中自由穿梭。于是,一个巨大的、透明的广袤空间--我们所认识的可见天空--出现了,它的半径约为4200万光年。因此,《创世纪》1:7-8中对天空的创造可以被解释为对宇宙历史中这一关键事件的引用。
下表总结了《圣经》中描述的宇宙创世过程和天文学的解释。比较结果表明,《创世记》中的创世叙述与天文学事实惊人地一致,这证实了上帝早已通过《圣经》揭示了这些真理,远在科学发现之前。
创世纪 | 天文学 |
真空涨落 (创1:2-创世之前) | 真空涨落 (大爆炸之前) |
创造光明 (创1:3-创世第一天) | 创造光明 (光子纪元) |
创造天空 (创1:7-8-创世记第2日) | 创造天空 (重组纪元) |
表1.1.《创世纪》中的创造与天文学的比较
c.地球和太阳哪个先被创造?
《创世纪》中创世第三天的主要事件是创造旱地和海洋。这可以理解为地球形成和构造的时期。水的聚集和旱地的出现标志着地球表面和地理特征的发展。《创世纪》第四天的主要事件是创造太阳。因此,地球是在太阳之前被创造的。研究《圣经》中的记载是否与天文观测相符一件有趣的事情。让我们来探讨一下。
恒星和行星都是由分子云形成的。分子云由约98%的气体(约70%的氢和28%的氦)和2%的尘埃(碳、氮、氧、铁等)组成。大部分恒星和类木行星都是由气体组成的,而大部分陆地行星都是由尘埃组成的。原恒星是分子云在自身引力作用下坍缩形成的。在这一过程中,分子云中的剩余物质形成一个旋转盘,称为原行星盘,这是行星最终成形的区域。引力坍缩引发核心的加热和压缩,导致原恒星的诞生,而周围的旋转盘则为行星体的形成和演化提供了环境。
随着原恒星继续收缩,它就变成了前主序星,并沿着赫兹普隆-拉塞尔图(H-R图)中被称为林轨(低质量恒星)和亨耶轨(高质量恒星)的恒星演化轨道运行。如果前主序的质量小于2个太阳质量,则可以观测到金牛座恒星;如果质量大于2个太阳质量,则可以观测到赫比格Ae/Be恒星。前主序星会继续收缩,直到其内部温度上升到1000万到2000万度。此时,前主序星开始进行氢核聚变,并成为天空中真正的恒星。处于这一阶段的恒星被称为主序星。
根据恒星演化理论和日震学研究,太阳在前主序星阶段停留了大约4000万到5000万年,之后就变成了主序星。
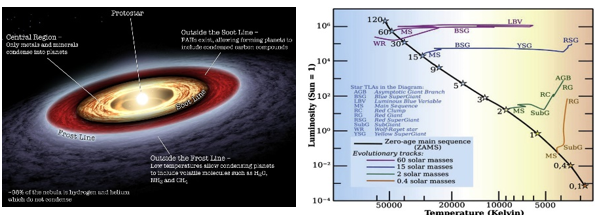
图1.10.原恒星和原行星盘,以及H-R图
当恒星在中心形成时,行星正在原行星盘中形成。尘埃粒子和气体碰撞形成鹅卵石,鹅卵石长成岩石,岩石长成行星碎片。这些行星体是行星的组成部分。
直到最近,人们才开始积极研究原行星盘中行星形成过程的细节。研究预测,从1毫米大小的鹅卵石形成一颗地球大小的行星需要几百万年的时间。这一预测可以用实际观测结果来检验,包括金牛座恒星HL Tau和PDS70的ALMA亚毫米波图像。
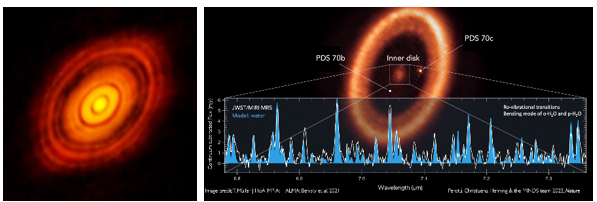
图1.11.HL Tau和PDS70
HL Tau的质量约为两个太阳质量,年龄约为一百万年。图像显示,几颗行星已经形成,并围绕着中央前主序星运行,原行星盘上的间隙就是证明。PDS70的质量约为0.76个太阳质量,年龄约为540万年。欧洲南方天文台VLT已经对两颗系外行星PDS70b和PDS70c进行了直接成像。2023年,詹姆斯-韦伯太空望远镜(James Webb Space Telescope)的光谱观测在原行星盘的类地行星形成区域探测到了水,并认为里面已经形成了两颗或更多的类地行星。值得注意的是,在HL Tau中看到的气体云和尘埃云在PDS70中被大部分移除,含有水的陆地行星已经在中心形成。
地球行星的形成需要540万年,但即使需要1000万年,也比太阳成为主序星的4000万至5000万年要短得多。这表明地球的诞生早于太阳,正如《创世纪》中所说的那样,并且与天文观测一致。
上帝在第三天所做的另一件大事是创造了植物和树木。无神论者和进化论者经常会问,如果太阳是在第四天被创造出来的,那么这些花草树木怎么可能存活下来呢?这个问题可以从恒星进化论的角度来解决。地球形成时,太阳还处于金牛座恒星阶段。虽然金牛座恒星不是主序星,但它们的表面温度在4000到5000开尔文之间。这种温度下的黑体辐射在可见光波段达到峰值。此外,太阳作为金牛座恒星时的大小是其目前大小的几倍。因此,它可以在可见光波长范围内提供足够的能量,使植物和树木能够进行光合作用。
d.地球的年龄是6000年吗?
“年轻地球创世论”认为地球和宇宙相对年轻,通常约有6000到10000年的历史,其依据是对《圣经-创世纪》中创世记的字面解释。年轻地球创世论者认为地球是在6个24-小时内被创造出来的,并拒绝接受有关地球和宇宙年龄的大部分现代科学共识。来自地质学、天文学和物理学等不同领域的大量科学证据表明,地球的年龄约为46亿年,宇宙的年龄约为138亿年。尽管证据确凿,但年轻地球创世论者并不认同。这种情况让人想起伽利略-伽利莱时代地心说和日心说之间的争论。
在进入主要讨论之前,让我们先看几个例子,让我们很容易理解地球和宇宙至少有几百万年的历史。
地壳由构造板块组成,这些板块缓慢移动,导致地震。没有人会否认这一事实。热点是岩浆从地壳下的地幔深处流出的一个点,其中心固定不动。当岩浆流出地壳并冷却后,就形成了陆地。夏威夷群岛就是这一过程的典型例子。在夏威夷大岛上,基拉韦厄火山仍然是一座活火山,当它喷发的岩浆在海水中冷却时,就会形成新的陆地。由于板块构造作用,新形成的陆地以每年约7-10厘米的速度向西北方向移动,这一过程造就了夏威夷的各个岛屿。这一过程造就了夏威夷的各个岛屿。即使是现在,这一过程仍在发生,这是不可否认的事实。
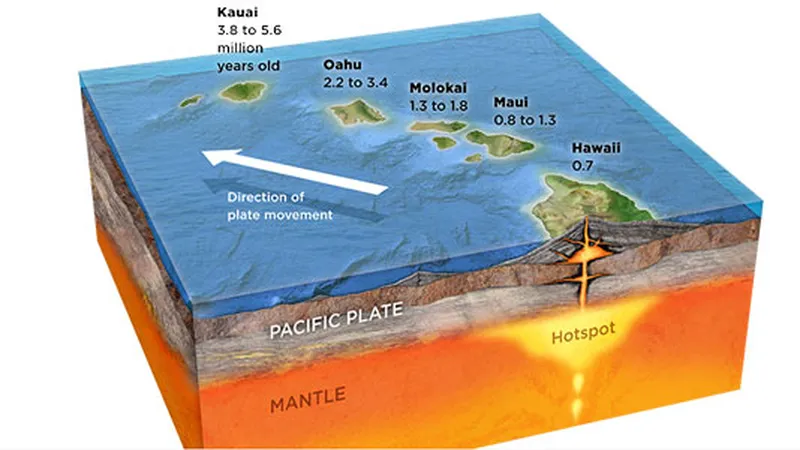
图1.12.夏威夷群岛的地质历史
考虑到地壳板块的移动速度,夏威夷群岛的年龄估计如下:大岛40万年,茂宜岛100万年,摩洛凯岛150万-200万年,欧胡岛(威基基所在位置)300万-400万年,可爱岛约500万年。在大岛,我们可以看到大部分的土地仍然覆盖着黑色的火山土壤,表明风化程度极低。与此相反,可爱岛经历了严重的风化作用,植被茂盛,因此被称为“花园岛”。这个例子提供了地球至少有几百万年历史的直接证据。
要直接理解宇宙至少有几百万年的历史,只需接受光的传播速度为每秒30万公里。太阳距离地球1.5亿千米。因此,我们现在接收到的阳光是8.3分钟前在太阳上产生的。太阳的体积大约是月球的400倍,但由于它距离地球更远,所以在天空中看起来和月球差不多大。没有人会否认这一点。仙女座星系的大小与我们的银河系相似,但距离我们有250万光年,因此它看起来是月球的四倍。我们能看到仙女座星系这一事实意味着,我们观测到的光是250万年前在仙女座产生的,现在才到达我们这里。如果你看过仙女座星系,你就无法否认这个事实。这是宇宙至少有几百万年历史的直接证据。
尽管有这些事实,但如果仍然坚持地球已有6000年的历史,就可能成为传播福音的绊脚石,而不是助手,有可能使许多人远离福音。因此,与其鼓吹年轻地球创世论,不如仔细阅读《圣经》中的《创世纪》,努力寻找解决之道,也许会更合理。
对于人类来说,时间总是从现在流向未来,而不会倒流。我们把一天定义为24小时,但如果我们被创造在其他星球上,一天就不是24小时了。例如,如果我们被创造在金星上,一天就是243个地球日,而在木星上,一天就是10个地球小时。因此,除非我们从地心角度改变对时间的定义和认识,否则很难解决这个问题。让我们带着这些事实进一步讨论这个问题。
i.《创世纪》中的日子
首先,让我们根据《创世纪》中的记录来估算宇宙的年龄。根据《创世纪》,上帝用六天时间创造了宇宙和其中的万物。根据《创世纪》5:3-32中的家谱记录,可以推算出从亚当到挪亚所经历的时间。诺亚的洪水发生在诺亚600岁的时候,从亚当到洪水的总年数是1656年。我们不知道诺亚的洪水是何时发生的。一些圣经学者和传统试图利用《圣经》中的家谱来确定洪水发生的时间,估计洪水发生在公元前2300-2400年左右。因此,根据这种解释,宇宙的年龄是7天+1656年+4400年=6056年。这就是年轻地球创世论者声称地球已有6000年历史的理论基础。
为了解决年代问题,让我们再来看看《创世纪》。虽然《创世纪》中的家谱记录似乎没有问题,但关于诺亚洪水的确切年份可能存在一些争论。然而,无论诺亚洪水发生在4400年前还是44000年前,都不会对138亿年的宇宙年龄产生重大影响。那么,解决日龄问题的关键在哪里呢?也许你已经注意到了--关键在于对创世头七天的解释。
原因很简单:“一天”的定义是我们所居住的地球的自转周期。要定义“一天”,太阳和地球都必须先存在。然而,《创世纪》,地球是在第三天被创造的,太阳是在第四天被创造的,但上帝在创造它们之前就使用了“昼”和“夜”这两个词。这意味着《创世纪》中的“昼”不是我们定义的24小时,而是上帝定义的“昼”。年轻地球创造论者的谬误在于他们误解了《创世纪》中提到的“日”是指人类字面意义上的24小时,从而导致了对《创世纪》中“日”一词的曲解。
如果《创世纪》中的日子不是人类定义的24小时,那么你可能会问:”《创世纪》中的日子按人类的日子计算有多长?虽然我们不知道确切的答案,但我们可以通过比较《创世纪》中描述的创世事件和宇宙大爆炸的创世事件来估算出一个大致的时间段。
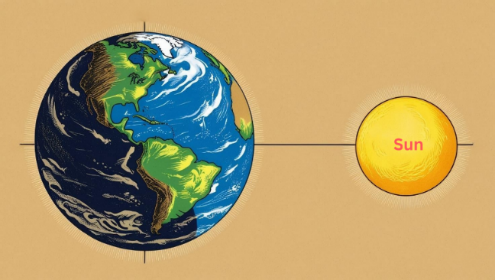
图1.13.要定义一天,地球和太阳必须事先存在。
创世第一天的主要事件是光的产生。宇宙大爆炸中的光子纪元与这一事件相对应,人类的创世第一天时间为38万年。创世第二天的主要事件是创造天空。重组纪元与此事件相对应,第二天的人类时间为10万年。第三天的主要事件是创造地球。如前所述,类地行星的形成大约需要1000万年,因此第三日的创世时间必定超过1000万年。第四日创世中最突出的事件是太阳的创造。由于太阳的形成需要4000万到5000万年,因此第四日的创世时间必定超过4000万年。下表总结了上述结果。
创造日 | 创世纪事件 | 天文学活动 | 人类时间 |
第一天 | 创造光明 | 在光子纪元创造光 | 38万年 |
第二天 | 创造天空 | 重组纪天空的形成 | 十万年 |
第三天 | 创造地球 | 创造地球 | > 1千万年 |
第四天 | 创造太阳 | 创造太阳 | > 4千万年 |
表1.2.用人类时间解释《创世纪》中的创世日
在这里,我们注意到一些关于上帝使用的时间概念的意想不到的事实。与人类一天24小时的时间相比,创世记中的日子要长得多。此外,上帝的时间并不是固定不变的,而是有变化的,从几十万年到几千万年不等。我们如何理解这一点呢?从某种意义上说,这不是一个令人惊讶的结果,而是一个意料之中的结果。
ii.时间的创造者
《创世纪》中使用的“日”在希伯来语中是yom(יום)。Yom”可以有多种解释,包括指年龄或很长一段时间。这种解释认为,创世的每一个“日”都代表一个漫长的时代,在此期间发生了具体的创世行为。另一种解释是,“yom”表示一段不确定的时间。这种观点认为,上帝的日子不受人类时间的限制,承认上帝作为时间的创造者,不受我们的时间限制。这种解释的例子可以在《圣经》中找到。
《新约-彼得后书》中写道
“亲爱的弟兄啊,有一件事你们不可忘记,就是主看一日如千年,千年如一日。”(彼得后书3:8)
这段话旨在鼓励那些耐心等待上帝应许的人。它还可能暗示,上帝对时间的看法与人类不同,这意味着上帝可以按照自己的意愿扩展或收缩时间。我们知道,时间不是一个固定的量。
根据狭义相对论,在同一惯性框架内,运动的观察者的时间比静止的观察者的时间走得慢(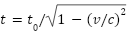 )。在广义相对论中,时间在强引力场中流逝得更慢(
)。在广义相对论中,时间在强引力场中流逝得更慢(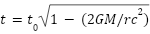 )。
)。

图1.14.时间膨胀示意图
上帝不仅能扩张或收缩,还能让时间停止。《旧约-约书亚记》中写道
“日头在天当中停住,不急速下落,约有一日之久。”(约书亚记10:13)
以赛亚先知求告耶和华,耶和华就使亚哈斯日晷上照下来的日影,往后退了十度。这个奇迹发生在约书亚与亚摩利人的战斗中,表明上帝有能力冻结时间。此外,上帝还创造了一个更加惊人的奇迹,《旧约-列王记下》中有这样的记载:
“以赛亚先知求告耶和华,耶和华就使亚哈斯的日晷,向前进的日影,退后了十度。”(列王记下20:11)
上面这节经文反映了上帝对希西家王含泪祈求长寿的回应。上帝怜悯希西家,赐给他15年的时间。为了证实祂的应许,神迹显现,使亚哈斯楼梯(日晷)上的影子向后移动了十步。这一神迹表明,上帝有能力逆转时间,而这一概念超出了我们目前的科学理解范围。

图1.15.亚哈斯的楼梯(日晷)
对于人类来说,时间是从现在到未来的单向流动,但对于上帝来说,正如《圣经》所示,时间是祂可以控制的变量。上帝可以缩短、延长、冻结,甚至扭转时间,这显示了他对自然规律的主权,也突出了人类的局限性和他的无限力量之间的对比。
e.微调宇宙
微调宇宙表达了这样一个事实,即构成和运行宇宙的基本物理常数都经过了极其精确的微调宇宙中才会有生命存在。
如果宇宙的密度大于临界密度,宇宙在形成后就会立即收缩。反之,如果宇宙密度小于临界密度,宇宙就会膨胀过快,阻碍恒星和星系的形成。无论哪种情况,我们都不会存在于这个世界上。
在《皇帝的新思维》一书中,彭罗斯使用贝肯斯坦-霍金黑洞熵公式来估算宇宙大爆炸时的几率。他计算出,宇宙以发展并支持我们所知的生命的方式出现的可能性是十分之一到十的幂次123。这表明,我们的宇宙并非产生于随机的机会或过程,而是通过神圣的造物主非凡的微调产生的!
引力常数、真空光速、普朗克常数、玻尔兹曼常数、电常数、基本电荷和微结构常数等物理学基本常数必须经过微调,宇宙中才能存在生命。如果这些常数稍有不同,宇宙就无法孕育生命。
例如,如果引力常数比现在小,引力就会减弱。引力减弱后,物质就不可能凝聚成恒星、星系和行星,包括我们今天生活的地球。如果普朗克常数比现在大,物理宇宙就会发生几个根本性的变化。首先,太阳辐射的强度会降低,导致从太阳到达地球的能量减少。能量的减少将影响许多自然过程,包括气候和天气模式。此外,普朗克常数值增大会增加原子的体积,因为原子能量水平的量化会发生变化。这种增加会削弱原子和分子的键合强度,从而降低化学反应的稳定性。植物依靠精确吸收光能将二氧化碳和水转化为葡萄糖的光合作用的效率将降低。依赖于当前量子力学平衡的整个生物化学和物理过程都将发生改变,从而导致生命环境的巨大差异和稳定性降低。
在基本常数中,精细结构常数引起了物理学家的特别关注。精细结构常数用希腊字母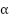 表示,它量化了基本带电粒子之间电磁相互作用的强度。
表示,它量化了基本带电粒子之间电磁相互作用的强度。
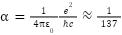
它是一个无量纲的量,近似值为1/137,这个数字自发现以来就一直吸引着物理学家。它的精确值对于宇宙的稳定和生命的存在至关重要。如果它与目前的数值哪怕有一点点,我们所知的生命就不会存在。
如果大于1/137,粒子之间的电磁相互作用就会变得更强。这将导致电子与原子核更紧密地结合在一起,缩小原子的体积,使重元素更容易形成,而氢等轻元素则更不容易形成。由于氢是核聚变的重要原料,这种变化会限制太阳和恒星产生能量所需的氢的供应,从而直接影响生命的生存。相反,如果小于1/137,粒子之间的电磁相互作用就会变弱。电子与原子核结合的紧密度会降低,从而导致原子和分子不稳定。这种不稳定性会导致原子和分子更容易衰变,从而阻碍像DNA和蛋白质这样的复杂分子的形成,而DNA和蛋白质是生命所必需的。因此,精细结构常数的任何重大变化都会对宇宙中物质的形成和生命的可能性产生深远的影响。
我们不知道其数值≈1/137的起源。狄拉克认为,的起源是“物理学中最基本的未决问题”。费曼将描述为“上帝之数”或“神奇之数”,它塑造了宇宙,而我们却不了解它。你可以说是“上帝之手”写下了这个数字,而“我们不知道他是如何下笔的”。如果我们重写的方程,它可以表示几个比值:电子的速度与光速的比值(即光的传播速度是电子的137倍)、静电斥力与单个光子的能量的比值、经典电子半径与电子的康普顿波长的还原比值。此外,电磁力与引力的强度之比为1036,电磁力与强力的强度之比为1/137。因此,无量纲常数的数值可以作为四种基本力的参考点。
如第三章《粒子物理与创世》所述,宇宙中所有物质(巴克伦)均由标准模型描述的基本粒子组成——夸克、轻子、规范玻色子和希格斯玻色子,总计17种。每种粒子都具有独特的质量、电荷和自旋。如果这些基本性质中的任何一个稍有不同,我们所知的原子、分子、生物和宇宙结构都将不复存在。
例如,如果上夸克和下夸克之间的质量差发生变化,维持质子稳定且中子仅略重于质子的微妙平衡将被打破。在这种情况下,氢无法形成,更重的原子核也无法合成,从而使原子无法存在。如果电子的质量有显著差异,原子大小和能级将发生变化,稳定的化学键将不再形成,从而阻止复杂分子的形成。如果希格斯玻色子的性质发生改变,赋予所有基本粒子质量的机制将被改变,从而重塑宇宙的结构。
此外,如果质子和电子的电荷不是完全相等且相反,中性原子就无法存在。如果夸克的电荷不同,质子和中子的性质将发生变化,从而破坏原子核的可能性。如果电子没有1/2的自旋,泡利不相容原理将不成立,原子就无法保持其结构。同样,如果玻色子没有整数自旋值,允许电磁力、强力和弱力等作用的量子场论框架将崩溃。最后,如果希格斯玻色子不是自旋为0的粒子,质量生成机制本身将失效,粒子将无法以当前形式存在。
精调的宇宙反映了支撑万物存在的惊人平衡与精确性。从宇宙的临界密度以难以想象的精确度设定,到彭罗斯对这种初始条件极小概率的计算,再到引力常数、普朗克常数和精细结构常数的微妙数值,每一个细节都指向一个为生命精确调校的宇宙。就连基本粒子本身——夸克、轻子、玻色子和希格斯粒子——也拥有恰到好处的质量、电荷和自旋,从而使原子、分子、恒星乃至最终的生命得以存在。这种和谐绝非盲目偶然所能解释。
这种非凡的精确性不仅令人惊叹,还迫使我们深入思考宇宙的起源与目的。物理定律的无缝互动带有刻意设计的痕迹,而神创论的概念则提供了一个深刻且令人信服的解释。正如管弦乐队只有在每件乐器都完美调音时才能奏出美妙的交响乐,宇宙也见证了造物主的智慧与力量,祂以目的与意义有序地安排了一切。
如果那些仅仅发现宇宙基本原理——万有引力、相对论、不确定性原理、泡利排斥原理和希格斯机制——的人被尊为天才并获得诺贝尔奖,那么创造者上帝又该有多么伟大?祂不仅构思了这些法则和原理,还创造了整个宇宙。
上帝的杰作 :地球
我们生活的地球为生物的生存提供了若干必不可少的微调条件。这些条件是如此精确,以至于它们常常成为微调宇宙的延伸
在此背景下,我们将探索地球的十种特殊条件,这些条件对于支持我们所知的生命而言尤为独特和关键。这些条件凸显了维持生物体所需的非凡平衡和精确性,使我们的星球成为浩瀚宇宙中的一片特殊绿洲。通过研究这些独特的属性,我们可以更深入地了解使生命在地球上繁衍生息的各种因素之间错综复杂的相互作用。
a.与太阳的正确距离
液态水的存在对生命至关重要。要想有液态水,行星必须在其中心恒星周围的特定区域内运行。如果行星离恒星太近,所有的水都会沸腾;如果行星离恒星太远,所有的水都会结冰。水既不会沸腾也不会结冰的轨道范围被称为“宜居带”。太阳系的宜居带估计在0.95AU和1.15AU之间(1AU是地球到太阳的距离)。因此,如果地球距离太阳再近5%或远15%,我们就不会在这里了。
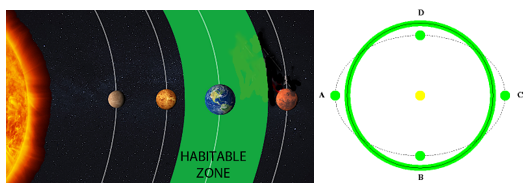
图2.1.太阳系中的宜居带(绿色)
在延伸到海王星(30AU)的黄道面上,宜居带所占的比例仅为0.05%。地球轨道的偏心率是影响宜居带范围的另一个重要因素。例如,如果偏心率大于0.5,那么所有的水都会在近日点附近每年沸腾两次,在远日点附近每年冻结两次。幸运的是,地球的偏心率只有0.017,因此轨道几乎是圆形的。
b.适度倾斜的地球自转轴
地球自转轴倾斜约23.5度。正因为如此,我们才能四季如春,气候温和。如果自转轴不倾斜(0度,参照水星的轴向倾斜=0.0度)或完全倾斜(90度,参照天王星的轴向倾斜=82.2度),会发生什么情况?
如果地球的自转轴不倾斜,气候、季节和宜居性将会发生一些重大变化。赤道将全年持续接受直射阳光,导致气温持续炎热。与此相反,两极将始终接受极少的阳光照射,导致长期寒冷。这种剧烈的温度反差将极大地影响全球气候和天气模式。
没有季节会对生态系统和农业产生深远影响。赤道附近的地区可能会变得过于炎热,许多农作物和生物都无法生长,而极地地区则依然寒冷刺骨。中纬度地区将成为主要的宜居区,但即使是这些地区也将缺乏许多动植物赖以维持生命周期和繁殖的季节变化。
人类社会将面临严峻的挑战,包括农业生产率下降和可居住地的压力增大。缺乏季节线索也会扰乱依赖季节变化的文化和经济活动。总之,一个不倾斜的地球将导致生命的活力减弱,环境变得更加恶劣。
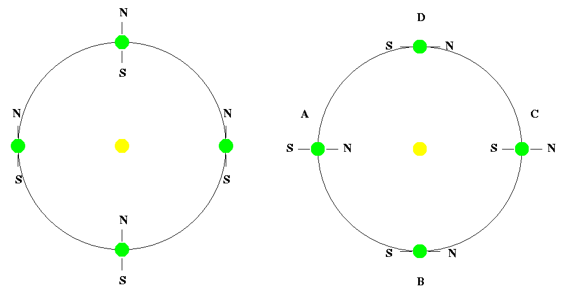
图2.2.地球的轴向倾斜。无倾斜(左)和90度倾斜(右)
如果地球自转轴完全倾斜到90度,将会对地球的气候和环境产生深远而巨大的影响。在这种情况下,一个半球将在一年中的一半时间里持续日照,而另一个半球将持续黑暗,然后在一年中的另一半时间里情况会发生逆转。
每个半球都将经历极端的季节变化。在夏季,一个半球会持续接受阳光照射,导致长时间的酷热和潜在的类似沙漠的条件。相反,在冬季,同一半球将经历持续的黑暗和严寒。
光照和温度的剧烈变化将严重破坏生态系统。许多动植物已经适应了当前的季节周期,这种极端变化将威胁到它们的生存。
依赖可预测季节的农业将受到严重影响。目前适合耕作的地区可能变得不适合居住,从而导致粮食短缺,农业生产方式也需要做出重大调整。
总之,完全倾斜的地轴会使地球更不适合生命的生存,造成极端和不稳定的环境条件。
c.拥有适当自转速度和公转速度的地球
地球的自转周期为24小时,其中昼夜各12小时。地球的自转周期决定了我们的生物节律。24小时的自转周期提供了8小时工作、8小时睡眠和8小时休闲的最佳时间段。然而,太阳系中并非所有行星都有最佳自转周期。例如,木星的公转周期约为10小时,而金星的公转周期为243天。
如果地球的自转周期缩短到10小时,将对地球的环境和生命产生重大影响。自转速度加快将导致昼夜缩短,造成白昼和黑夜的快速交替。这会扰乱许多生物的昼夜节律,影响睡眠模式、进食行为和繁殖周期。
自转速度的增加还将导致更强的科里奥利效应,加剧天气模式,并可能造成更严重的风暴和飓风。更快的自转还可能影响地球的构造活动。离心力的增加可能会导致更频繁、更剧烈的地震和火山爆发。
另一方面,如果地球的自转周期与金星一样为243天,那么地球及其居民将面临严重的后果。如此缓慢的自转将意味着极长的昼夜,每个昼夜大约持续120天。
朝向太阳的一侧会经历长时间的加热,导致炙热的温度,而朝向太阳的一侧则会经历长时间的黑暗和严重的冷却,可能会结冰。这些极端温度将使大多数生命形式的生存面临挑战。长时间的升温和降温会破坏大气环流,可能导致极端天气模式。飓风、大风暴、长期干旱或洪水可能成为常见现象。
长时间的日光和黑暗会严重扰乱动植物的生命周期,影响光合作用、繁殖和进食模式。
人类活动、农业和基础设施需要进行重大调整,以应对恶劣和多变的条件,这对生存和日常生活构成了巨大挑战。
地球的轨道周期对人类的生存也很重要。地球的轨道周期为365天,春、夏、秋、冬各3个月。每个季节的长度都很均衡,确保没有一个季节太短或太长。这种平衡对于农业周期、植物生长、动物迁徙时间以及其他生态过程都至关重要。
如果地球的轨道周期很短,比如88天,类似于水星,会发生什么情况呢?在这种情况下,每个季节只持续3周左右。地球上的大多数农作物从春天播种到秋天收获需要6到9个月的时间。然而,如果每3周换一次季,农作物就没有足够的时间成熟,从而导致严重的粮食短缺,直接影响人类的生存。
相反,如果地球的轨道周期很长,比如164年,类似于海王星,会发生什么呢?每个季节将持续约40年。夏季的延长会导致热浪和潜在的沙漠化,而冬季的延长则会造成长时间的寒冷和冰冻,影响农业和生态系统。虽然人类可能会通过适应来避免食物短缺,但野生动物却会在长达40年的冬季里艰难地寻找食物。长期的恶劣条件将使大多数野生动物几乎无法生存,导致大面积灭绝。
d.合适的尺寸
你可能没有想到,地球的大小对人类的生存至关重要。地球的大小影响着它的引力,而引力又影响着从维持生命的大气层到支持稳定的水体和维持保护性磁场的能力等方方面面。
如果地球只有现在的一半大小,重力将减小到现在的一半。重力的减少将对地球支持生命的能力产生重大的、潜在的破坏性影响。减少的重力可能不足以保留浓密的大气层。更稀薄的大气层将减少对有害太阳辐射和流星体的保护,也可能无法支持生命所需的稳定天气模式。
重力减小还会影响液态水的保留,导致蒸发率增加,地表水可能会随着时间的推移而流失。这将使海洋、河流和湖泊难以为继,而它们对于支持多样化的生态系统和人类文明至关重要。
此外,地球变小后,磁场会减弱,对太阳风的保护作用也会减弱。这可能会剥离大气层,使地球表面进一步暴露在有害的宇宙和太阳辐射下,从而使地球更不适合人类和其他形式的生命生存。
图2.3.太阳系行星大小比较
如果地球的大小是现在的两倍,对重力和逃逸速度的影响将是巨大的,并对地球上的生命产生深远的影响。重力会增加,使地球上的一切都感觉更重,逃逸速度也会增加一倍。重力的增加会使人类和其他生物的运动更加吃力,随着时间的推移,可能会导致更大的身体压力和适应性。
引力和逃逸速度的增加也会对大气层产生影响。更强的引力会保留更多的气体,包括甲烷和氨等有毒气体,类似于土星和木星的大气层。这些气体可能会积累到有害的程度,形成一个不适合大多数生命形式的有毒环境。
此外,重力增加可能会影响地质过程,导致火山活动更加剧烈,山脉更高。总之,地球面积增大,引力和逃逸速度增加,将给生命的生存带来巨大挑战,可能导致环境更加恶劣和不稳定。
e.磁层的存在
地球周围有一个被称为磁层的磁场系统,它能保护地球免受有害的太阳和宇宙辐射。这个保护罩对于维持地球上的生命至关重要。要拥有磁层,两个因素必不可少:适当的自转速度和金属液体外核的存在。幸运的是,地球拥有这两个因素。行星的自转在液态外核中引起流体运动(对流),产生强磁场,形成磁层。
如果我们没有磁层会怎样?如果地球没有磁层,生物体和大气层将面临严重的后果。如果没有这个保护罩,有害的太阳辐射和宇宙辐射将轰击地球,大大增加生物体患癌症和基因突变的风险。此外,磁层通过偏转太阳风中的带电粒子来帮助防止大气流失。如果没有磁层,随着时间的推移,这些粒子将通过溅射过程带走大气层,耗尽氧气和氮气等必需气体。这种大气侵蚀会导致大气层变薄、地表压力降低、温度变化剧烈,从而使地球不再适合生命生存。
火星上的磁场强度约为地球的0.01%。由于磁场较弱,火星上无法形成全球磁层,因此大部分空气被溅射过程带走。
磁层的磁场线在北极和南极附近的两极汇聚,导致磁场强度自然减弱。这会导致这些地区受到的太阳辐射增加。高能带电粒子电离并激发高层大气中的原子,产生五彩缤纷的北极光和南极光。
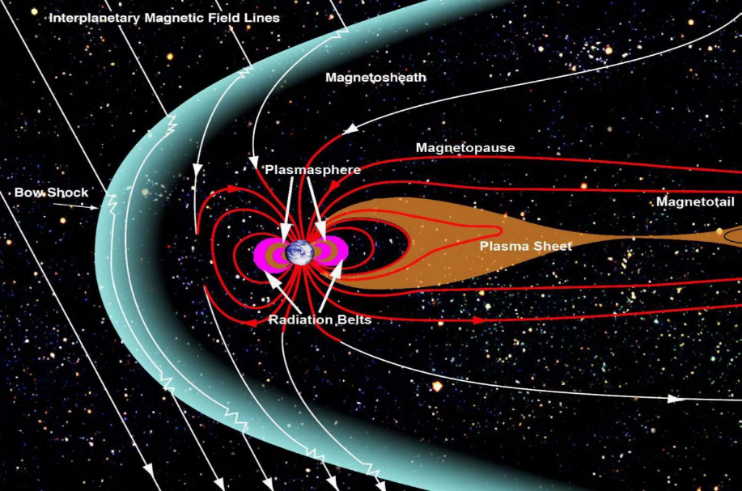
图2.4.地球磁层偏转有害宇宙射线
f.特大月球的存在
月球对地球生命体的存在具有重要影响。虽然我们常在夜空中仰望的月亮与人类生存看似毫无关联,但它实际承担着两大关键作用:其一是稳定地球自转轴以防止气候异常,其二是通过潮汐力维持海洋生态系统的平衡。
首先让我们了解月球对地球自转轴的稳定作用。正如前文所述,地球自转轴倾斜23.5度,为地表生态系统带来了宜居的季节更替。而地球自转轴能保持这种恒定的23.5度倾角,正是由于近在咫尺的月球存在。
在太阳系的类地行星中,没有哪颗行星拥有像地球这样巨大的卫星(半径=1740公里)。同为类地行星的水星和金星根本没有卫星,火星虽有两颗以希腊神话中福波斯和戴摩斯命名的卫星,但其半径仅约10公里,相当于月球半径的0.6%,极其微小。
如前所述,太阳系中适宜生命生存的宜居带位于1天文单位附近。而太阳系行星运行的黄道面延伸至海王星附近约30天文单位。考虑到这一距离,地球实际上过于贴近太阳。唯有如此靠近才能维持适宜生存的温度,但存在一个问题:太阳对地球的引力过于强大,导致地球自转轴不稳定。
太阳引力引发的自转轴摆动现象在邻近行星中亦可观测到。下图展示了火星(因缺乏大型卫星)在太阳引力作用下自转轴与离心率的变化规律。图中可见火星自转轴与离心率以约15万年为周期剧烈振荡:过去600万年间,自转轴倾角从15度变至45度,离心率从0.01变化至0.11。
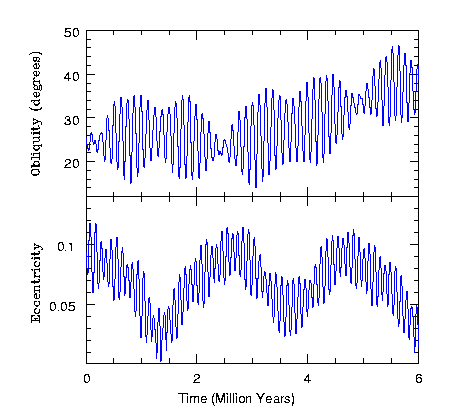
图2.5.的自转轴和偏心率变化
与地球大小相近的姐妹行星金星因缺乏卫星,其自转轴受太阳引力影响会发生0度至180度的混沌变化。若假设地球没有卫星,其自转轴可能产生怎样的变化?不同论文观点虽有差异,但普遍认为数百万年间自转轴将发生约0度至70度的剧烈摆动。如此剧烈的变化将引发严重的气象灾害,最终可能对生命体生存造成致命打击。所幸造物主为地球预备了这颗异常巨大的卫星,从而避免了灾难发生。
那么,地球为何能拥有如此巨大的卫星,而其他类地行星却未能如法炮制?关于月球起源,学界提出过多种假说:撞击说认为火星大小的天体与地球相撞形成月球;捕获说主张月球是被地球引力捕获的其他行星;同步形成说主张月球与地球同时诞生;分裂说则认为月球是地球部分物质脱离形成的。然而最新研究倾向于支持撞击说作为最合理的解释。但要通过巨型撞击形成地球-月球系统这类大型卫星,必须满足多个精密条件:
首先,撞击体的大小与撞击角度必须高度契合。撞击体(泰亚)的质量需约为地球的10%,且必须以倾斜角度撞击——既不能完全摧毁地球,又需将地幔物质充分喷射至太空。正面碰撞无法产生形成月球的物质。
其次,喷射出的残骸物质必须快速凝结并聚合。在残骸被地球强大的潮汐力分散或重新吸回之前,必须在短暂的聚合窗口期内完成月球形成。
关键在于,月球稳定形成的必要条件是撞击后地球自转速度足够快。因为只有如此,撞击喷射出的物质才能避免重返地球,在罗什极限外形成稳定轨道并聚集成月球。正因如此,当大小、角度、速度与时间条件完美契合时,其他类地行星形成如此巨大卫星的现象,据推测应极为罕见。
海潮主要是由月球引力引起的。潮汐为漂浮的浮游生物提供氧气,并将它们分散到广阔的区域,被小鱼吃掉。潮汐还将营养丰富的淡水与海水混合,将这些营养物质输送给浮游生物和小鱼。如果没有潮汐,营养丰富的淡水就不会与海水混合,从而导致无法控制的藻类大量繁殖。如果藻类含有毒素,这些藻华就会产生赤潮或有害藻华(HABs),可导致鱼类、海鸟、哺乳动物甚至人类死亡。即使藻类无毒,它们也会在腐烂过程中消耗掉水中所有的氧气,堵塞鱼类和其他海洋生物的鳃。如果没有月球,海洋生态系统早就被破坏了。此外,我们也不会有海鲜,包括龙虾、虾和寿司。
不过,即使地球上的月球比现在的大小更小或更大,或者月球的位置比现在的位置更远或更,我们仍然可能面临类似的问题。

图2.6.赤潮
g.地球守护神木星的存在
木星是太阳系中最大的行星,比地球大11.2倍,重318倍。木星的存在对我们的生存非常重要。地球不断受到陨石(主要是碎裂的小行星和彗星碎片)的轰击。陨石坠落的频率为每小时一次一米大小的陨石,每天一次几米大小的陨石,每年一次几米到十米大小的陨石,每十年一次十几米大小的陨石,每百年一次十几米到一百米大小的陨石。
当小于10米的陨石进入大气层时,大部分会因大气摩擦和压缩而燃烧。但是,如果陨石大于10米,就会发生灾难性事件。1908年,一颗约55米大小的陨石在通古斯卡地区上空5至10公里处爆炸,摧毁了约8000万棵树木,影响范围达2150平方公里。
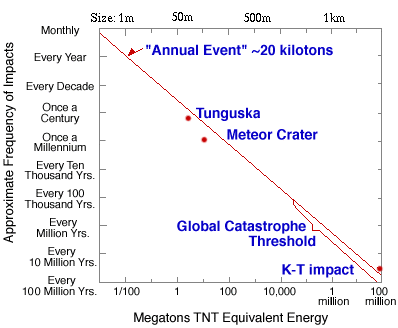
图2.7.落在地球上的陨石的大小和频率

图2.8.掉落在通古斯卡的陨石砸倒的树木
木星之所以至关重要,是因为它就像一个宇宙吸尘器,可以捕捉陨石和彗星,否则这些陨石和彗星可能会撞击地球,造成像通古斯卡事件那样的灾难性事件。模拟结果表明,木星捕捉彗星的效率比地球高出约5000倍。1994年,木星捕获了碎裂的舒梅克-列维9号彗星,这就是一个显著的证明。如果这颗彗星撞击地球,它可能会将尘埃和碎片送入大气层,阻挡阳光。这种阻挡可能持续很长时间,足以杀死所有植物,导致依赖植物生存的人类和动物灭绝。

图2.9.碎裂的舒梅克-列维9号及其对木星的撞击
h.板块构造的存在
板块构造学是一种描述地球岩石圈大尺度运动的理论,地球岩石圈在地幔对流运动的作用下分裂成几个大的构造板块。这一理论解释了许多地质现象,包括大陆的移动、山脉的形成、地震和火山活动。
图2.10.构成地壳的板块
板块构造在地球系统的各个方面发挥着至关重要的作用,直接或间接地影响着人类的生存。板块构造最重要的方面之一是通过碳循环自动调节地球气候。
地球气候主要由太阳辐射、地球表面反照率和大气成分决定。其中,入射太阳辐射在很长一段时间内几乎是恒定的。反照率是入射辐射与反射辐射的比值。来自地球表面的反射辐射有很大一部分会被大气中的二氧化碳(CO2)分子所吸收。被吸收的辐射会加热CO2分子并将其重新辐射到各个方向,其中约有一半会以热能的形式返回地球。这种被截留的热能会增加全球平均地表温度,这就是所谓的温室效应。
碳循环是碳在大气、海洋、土壤、矿物、岩石、植物和动物之间进行交换的过程,对调节地球气候至关重要。碳以CO2的形式从呼吸、燃烧和火山爆发中进入大气。植物在光合作用中吸收CO2,将其转化为有机物,有机物被动物消耗,并通过呼吸和分解释放回大气中。在海洋中,CO2被海洋生物溶解和利用,形成碳酸钙(CaCO3)外壳。当这些生物死亡时,它们的外壳会堆积在海底,形成沉积岩。
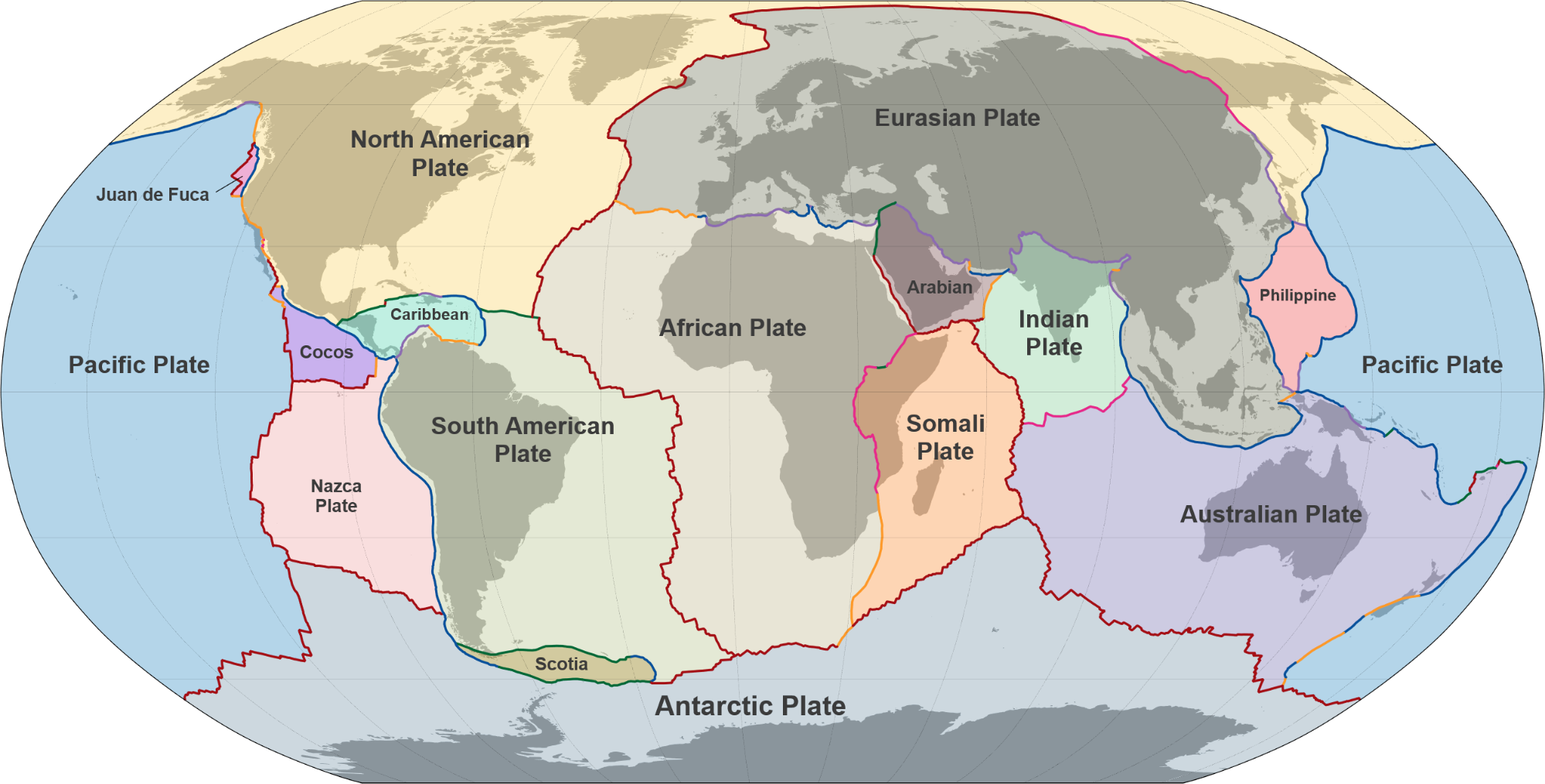
陆地上的岩石风化也会吸收CO₂,形成碳酸盐被冲入海洋。这一风化过程取决于温度。如果大气中的CO2过多,温室效应导致温度升高,那么风化过程就会增加,吸收更多的CO2。如果大气中的CO2被清除,那么地球的温度就会下降。如果地球温度降低,风化过程就会减弱,从大气中清除的CO2就会减少。如果出现这种情况,那么累积的CO2就会产生更多的温室效应,使温度升高。这一过程被称为“二氧化碳岩石风化循环”。在地质时间尺度上,构造活动会通过俯冲作用将这些富含碳的岩石推入地幔。然后,碳通过火山爆发释放回大气中,完成循环。与温度相关的二氧化碳岩石风化循环自动调节着地球在地质时间尺度上的温度。下图显示了这一循环在过去80万年中的运行情况:当二氧化碳量增加时,地球温度升高;当二氧化碳量减少时,地球温度降低。
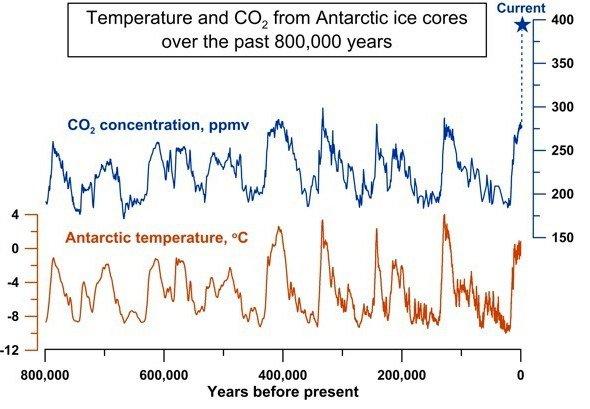
图2.11.CO2与温度的相关性
然而,如果没有板块构造作用,二氧化碳岩石风化循环就不会起作用。在这种情况下,积累的二氧化碳将无法循环利用,因此温室效应也会减弱。如果没有温室效应,地球的温度就会迅速下降,所有的水域都会冻结。如果所有水域都被冻结,由于反照率大,传入的太阳能将被反射,最终地球将进入不可逆转的冰河时期。
有关板块构造的最新研究表明,如果地球比现在大或小20%,如果地壳中铁和镍等金属含量稍多一些,或者如果地壳更厚一些,板块构造就不会像现在这样发挥作用。
总之,板块构造是一个基本过程,它通过维持地球地质和环境的稳定来支持生命。
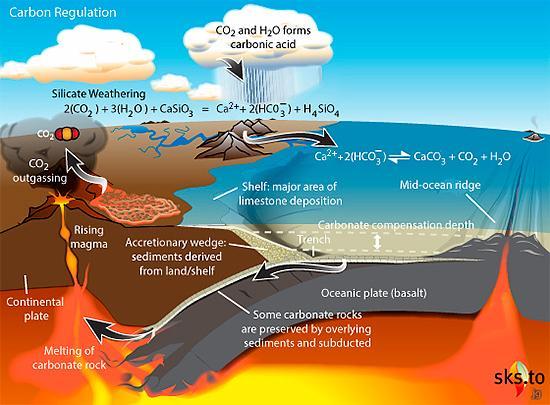
图2.12.板块构造对二氧化碳的循环利用
i.太阳的正确大小
行星宜居带(HZ)的大小取决于其中心恒星的大小和类型。
对于红矮星等小恒星来说,HZ离恒星很近,因为恒星发出的光和热较少。这使得可居住区的范围比围绕太阳的范围更窄。由于距离较近,红矮星宜居带中的行星可能会被潮汐锁定,就像月球对地球一样。如果发生这种情况,由于行星自转缓慢,它将无法产生磁场并形成磁层。如果没有磁层,来自恒星的有害辐射就会随意到达行星表面,损害细胞和DNA。此外,白昼一侧将经历持续的日光和酷热,而黑夜一侧将永远处于黑暗和极寒之中。
对于蓝巨星或红巨星等大型恒星来说,HZ距离恒星更远。然而,位于这些区域的行星面临着巨大的挑战。巨星由于质量大而演化迅速,很快就会烧尽氢气,膨胀为红超巨星,并经历多个阶段的核聚变,直至形成铁质内核。铁核最终坍缩,导致超新星爆炸,留下中子星或黑洞。巨星的典型寿命只有几百万年,这意味着在恒星爆炸成为超新星之前,其HZ内的任何行星居民都需要找到另一颗合适的行星迁移到那里生存。此外,巨星会发出大量的紫外线和X射线辐射,这些辐射会对DNA和细胞造成伤害,从而使HZ内行星的表面环境不再那么适合生命生存。此外,巨星在能量输出方面会表现出,导致环绕行星的气候不稳定。这种不稳定会导致极端的温度波动,使生命难以生存。
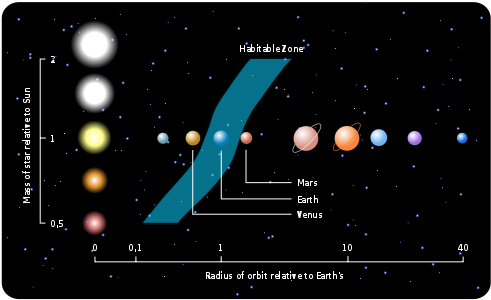
图2.13.宜居带随恒星变化
类太阳恒星周围的宜居带(HZ)具有许多优势。这些恒星具有长期相对稳定的能量输出,为其宜居带中的行星提供稳定的光和热。这种稳定性有助于形成稳定的气候和生态系统。类太阳恒星周围的宜居带距离适中,既不会离恒星太近,也不会太远。类太阳恒星的光谱非常适合光合作用,使植物和其他光合生物能够有效地将阳光转化为能量,形成可持续食物链的基础。此外,与红矮星等较小的恒星相比,类太阳恒星通常具有较低水平的有害恒星活动。较少的耀斑和较低强度的磁活动意味着宜居带内的行星较少暴露于潜在的、破坏性辐射和大气剥离。
类似太阳的恒星只占百分之几,因为大多数恒星都比太阳小,比太阳轻。太阳是一颗恒星,但大约50%到60%的恒星是双星或多恒星系统。由于复杂的轨道、多变的光照、引力扰动和潜在的辐射水平,多恒星系统的宜居带受到的限制要大得多。
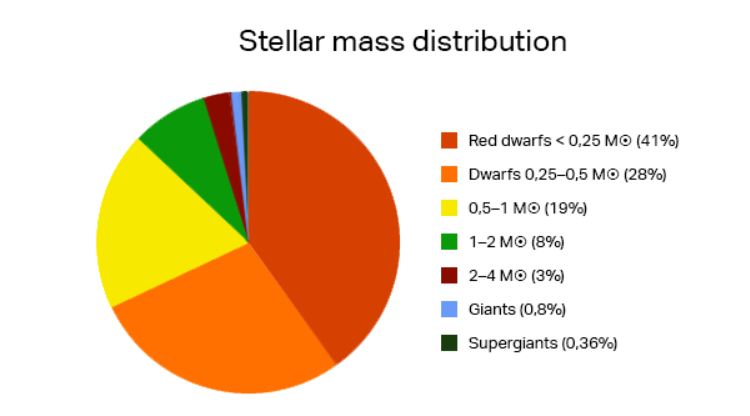
图2.14.恒星的质量分布
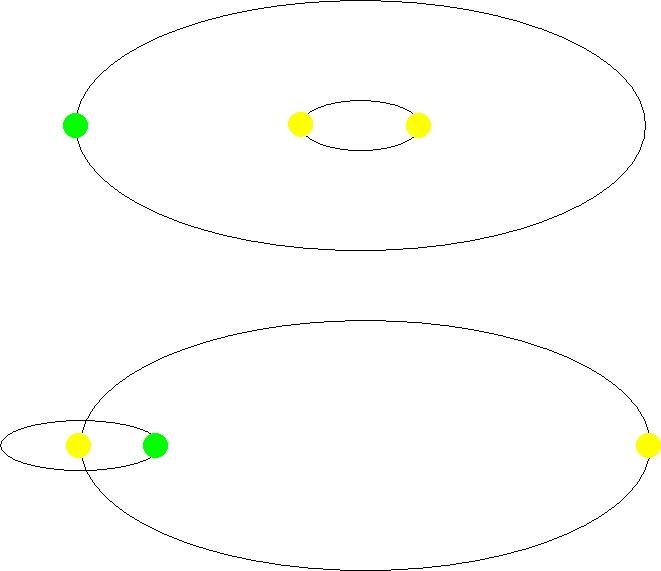
图2.15.双星系统中的环双星轨道(左)和环主轨道或环次级轨道(右)
j.银河系中心的正确距离
就像太阳系中的宜居带一样,银河系中也存在一个银河宜居带(GHZ),那里的条件对生命最为有利。银河宜居带所需的条件包括金属性、恒星密度、辐射水平和轨道环境。
GHZ需要有形成陆地行星和有机分子所必需的重元素(比氦重的元素)的最佳浓度。虽然银河系中心的金属元素更为丰富,但由于恒星密度较高,导致超新星爆炸、伽马射线暴(GRB)和其他高能事件频发,因此这一区域并不能被视为GHZ的有利区域。
发生在距离地球1万光年范围内的伽马射线暴可能会对地球的大气层、气候和生物圈造成破坏性影响。直接影响包括由于臭氧层遭到约40%的破坏而导致紫外线辐射增加,而长期影响则可能包括显著的气候变化和大规模物种灭绝。这种事件将对人类文明和自然界构成严重威胁。40%的臭氧层遭到破坏后,增加的紫外线辐射对DNA的破坏程度将增加16倍。
图2.16.浮游植物
浮游植物是海洋食物网的基础,对紫外线辐射特别敏感。紫外线照射的增加会抑制它们的生长和繁殖,导致浮游植物数量减少。浮游植物在光合作用中吸收CO2,在碳循环中发挥着至关重要的作用。浮游植物的减少会降低这种碳固存作用,可能会加剧大气中CO2的积累,加剧温室效应。
有证据表明,地球上过去发生的大灭绝事件可能是由附近的全球热辐射光束引发的。例如,一些科学家推测,4.5亿年前的奥陶纪-志留纪生物大灭绝事件可能是受到了距离地球6000光年的一个全球热辐射光束的影响。
银河系中心遇到的另一个问题是经常与其他恒星近距离相遇。这些近距离相遇会造成巨大的引力扰动,从而破坏行星系内行星轨道和旋转轴的稳定性。这种扰动可能导致轨道交叉、碰撞或抛出系统。附近恒星的引力影响也可能扰乱奥尔特云和柯伊伯带中天体的轨道,将更多的彗星和小行星送入内太阳系。这将增加撞击包括地球在内的行星的可能性。
银河系外围的恒星密度较低,不存在这些问题,但有一个关键问题:超新星爆炸率低。这就导致星际介质中缺乏足够的金属元素来形成陆地行星,从而使银河系外围不利于GHZ的形成。
在GHZ的有利区域,有足够的重元素形成行星,较少的超新星和其他危险事件为生命提供了安全的环境,较少的拥挤区域为稳定的行星轨道提供了条件。此外,在恒星的轨道速度与银河旋臂的模式速度相匹配的区域,即冠状半径。在冠状半径范围内,恒星及其行星系统与旋臂发生的破坏性引力相互作用较少,从而提高了持续宜居条件的可能性。
考虑到所有这些条件,GHZ位于距银河系中心23000到29000光年之间。巧合的是,我们的太阳系距离银河系中心26000光年,位于GHZ的中心。

图2.17.银河系中的银河宜居带
在本章中,我们探讨了使地球成为一颗非凡行星的十种独特而非凡的条件。这些条件是如此错综复杂的平衡和精确的校准,以至于它们随机发生的可能性低到了天文数字。地球与太阳的距离、轴向倾角、自转周期、磁场、大气和其他关键因素的精确性,创造了一个能够支持生命的独特环境。这种有利条件的组合在宇宙其他地方同时出现是非常不可能的,这进一步凸显了地球的独特性。此外,地球所享有的保护和稳定性--免受有害宇宙事件的影响和保持微妙的生态平衡--也进一步证明了它在其他行星中的独特性。这些因素共同有力地支持了这样一种观点,即地球是由神圣的造物主有意设计为生命栖息地的。这种微调的平衡条件不仅仅是一种巧合,而是一种有目的的智慧设计,使地球成为一个非同寻常、独一无二的生命生存环境。
创造还是进化?
我们是被创造的还是进化来的?关于生命起源的争论仍在继续,但目前的教育体系将进化论作为有关生命起源的既定理论来教授,而将创造论视为不科学的主张。
进化论以“生物起源”假说为起点来解释生命的起源。我们将首先详细探讨这一问题,然后探讨达尔文的理论应被称为“进化论”还是“遗传适应论”。我们还将讨论人类是否由猿进化而来的问题。此外,我们还将介绍智能设计,并从粒子物理学、外星生命的存在、动物本能和自然界中的数学角度来审视创世论。
a.生命的起源
关于创造论与进化论有效性的全部争论,最终都取决于一个核心问题:地球上最初的生命是如何出现的?如果生命纯粹通过自然的、无指导的过程产生,那么进化论就得到了根本性的验证。反之,如果生命在没有造物主干预的情况下不可能产生,那么创造论仍是唯一有效的解释。
所有生物体皆由细胞构成,而每个细胞都由核酸(DNA和RNA)、蛋白质、脂质和碳水化合物等主要大分子构成。作为主要功能分子的蛋白质,是由氨基酸长链组成的,其序列由DNA中存储的遗传信息决定。此外,脂质和碳水化合物是由遵循DNA编码指令的酶合成的——而这些酶本身就是蛋白质。
因此,即便构建最简单的细胞,蛋白质也绝对不可或缺;然而要产生蛋白质,DNA和RNA必须已然存在。但在原始地球上,这些物质——DNA、RNA、蛋白质或酶——皆不存在。由于这种循环依赖关系,生命自然起源的假说从一开始就注定无法成立。
然而,进化论者常坚称在早期地球上,RNA曾以某种方式自行形成。这一被称为“RNA世界假说”的理论认为,RNA同时兼具两种功能:既作为遗传分子(如mRNA)存储信息,又作为催化酶(核酶,如rRNA)合成蛋白质。
然而现实中,单个功能性RNA分子需要极其复杂精确的核苷酸序列,其自然随机形成的可能性近乎为零。即便存在极低概率的偶然出现,此类RNA分子也因化学性质极不稳定而在水中迅速分解。此外,已知核酶的催化能力极其有限,远逊于真正的蛋白质酶。最根本的矛盾在于:没有蛋白质就无法高效复制RNA,而没有RNA又无法合成蛋白质。因此,RNA世界假说存在着严重的内在矛盾。
此外,即便假设RNA、蛋白质和DNA都偶然形成,后续步骤——自组装成原核细胞、进化为真核细胞、细胞器定位、细胞分化、组织器官形成,直至多细胞生物的出现——每一步都代表着一系列概率微乎其微的事件。因此,本章将严谨探讨生命是否真能仅凭随机过程产生。
我们将首先重温经典的米勒-尤里实验,该实验常被用于探讨生命化学起源。我们将评估其实验结果及其公认的局限性——该实验仅试图证明在假设的早期地球条件下,无机前体物质可形成简单有机化合物。随后我们将逐一探究以下课题:i)氨基酸形成,ii)RNA形成,iii)蛋白质形成,iv)DNA形成,v)原核细胞形成,vi)真核细胞形成,vii)细胞器定位,viii)细胞分化,ix)组织器官形成,x)多细胞生物形成。通过这种系统分析,我们将确定进化论所宣称的每个阶段——从非生命到生命——是否真的可能通过随机、无指导的过程发生。
i.米勒-尤里实验
米勒-尤里实验是探讨生命化学起源时最常被引用的研究之一。该实验旨在验证:在模拟早期地球环境的条件下,无机物质能否自然生成简单有机分子——尤其是作为蛋白质基本单元的氨基酸。为实施该实验,米勒与尤里假设原始大气由甲烷、氨、氢和水蒸气构成。他们将这种气体混合物置于封闭装置中循环,该装置配备加热单元、真空泵、模拟闪电的放电装置及冷却系统。运行数日后,他们报告在反应产物中检测到四种氨基酸。
尽管该实验确实证明在受控实验室条件下,简单气体可形成少量氨基酸,但其实际与早期地球大气环境的关联性及实验装置的适用性仍存争议。基于这些原因,米勒-尤里实验值得重新审慎评估。
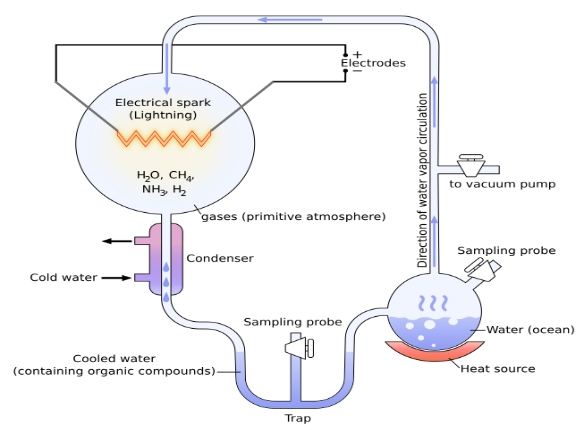
图3.1.米勒-乌雷实验示意图
米勒-乌雷实验使用放电装置来模拟自然闪电,但他们的装置与自然闪电在许多方面有很大不同。他们的装置使用的电压为5万伏,产生的热量为250度,而闪电的电压为1亿伏,产生的热量为5万度。米勒-乌雷实验中的放电是相对连续的,可以持续很长时间,从而确保化学反应有稳定的能量输入。相比之下,闪电并不是连续发生的,而是零星出现,而且持续时间极短,只有几微秒到几毫秒。
彗星是早期太阳系的残余物,含有相对保持不变的原始建筑材料。彗星的成分可以为了解早期地球大气层的成分提供宝贵的信息。彗星的主要成分是水(86%)、二氧化碳(10%)和一氧化碳(2.6%)。氨和甲烷各占不到1%。这一结果表明,米勒-乌雷实验中使用的气体并不能准确地代表早期地球的大气层,因为它不包含含量最高的二氧化碳和含量第二高的一氧化碳。此外,二氧化碳是一种氧化剂,会抑制氨基酸的形成。
组成 | 比率(%) | 参考资料 |
水(H2O) | 100(86%) | Pinto等人(2022年) |
二氧化碳(CO2) | 12(10%) | Pinto等人(2022年) |
一氧化碳(CO) | 3(2.6%) | Pinto等人(2022年) |
氨气(NH3) | 0.8(0.7%) | Russo等(2016) |
甲烷(CH4) | 0.7(0.6%) | Mumma等人(1996年) |
表3.1.彗星的成分(水=100)
米勒-乌雷实验假定早期地球的前生物大气层是还原性大气层。然而,如果它是氧化性大气,就会通过分解或氧化有机分子而阻碍氨基酸的形成。早期地球大气层的条件是科学界一直在探索和争论的问题。Urey(1952年)、Miller(1953年)和Chyba&Sagan(1997年)认为大气层是还原性的,而Albeson(1966年)、Pinto等人(1980年)、Zahnle(1986年)和Trail等人(2011年)则认为大气层是氧化性的。
值得一提的是Trail等人(2011年)在《自然》杂志上发表的论文。他们利用铈(Ce)氧化态的比率分析了哈代锆石晶体的氧化态。分析结果表明,哈代岩浆的氧化程度比以前想象的要高,其条件与现代火山气体的条件类似。哈代岩浆的氧化态更强意味着火山,释放出的氢气(H2)更少,而水蒸气(H2O)、二氧化碳(CO2)和二氧化硫(SO2)更多。他们的结论是,早期地球大气的还原性可能比传统认为的要低,而氧化性要高。他们的研究结果对米勒-乌雷实验的有效性提出了质疑,认为在前生物早期地球上可能无法通过生物生成形成氨基酸。
实验中产生的氨基酸是在实验室条件下收集和保存的。在地球早期恶劣多变的条件下,这些化合物可能不太稳定,更容易降解。实验中有机分子的浓度受到控制,并保持在相对较高的水平。而在早期地球上,这些分子可能会在广阔的海洋中被高度稀释或迅速分散,从而可能减少进一步化学进化的机会。
另一个关键问题是手性。产生的氨基酸是外消旋的,这意味着它们含有等量的左旋和右旋异构体。地球上的生命主要使用左旋氨基酸(99.3%),米勒-乌雷实验仍无法解释这种同手性的起源。此外,在模拟原始地球环境下进行的实验从未同时产生过全部20种L型氨基酸。即便所有氨基酸都偶然形成,它们聚合为精确序列以构成功能性蛋白质——或自发组装成RNA和DNA等核酸的概率在统计学上微乎其微。
基于此,我们现在转向一个关键问题:被视为生命起源关键成分的RNA,在原始地球环境中是否可能自然形成?抑或这种情景实际上根本不可能发生?
ii.RNA的形成
要理解RNA在自然界中形成是何等困难,我们首先必须了解RNA的实际构成。RNA是一种由数百至数千个核苷酸通过磷酸二酯键连接而成的长链聚合物。每个核苷酸又由三部分构成:含氮碱基(腺嘌呤A、鸟嘌呤G、胞嘧啶C或尿嘧啶U)、五碳糖核糖,以及一个或多个磷酸基团。
在原始地球上,即使要自发形成单个功能性RNA分子,这三种成分都必须在生命起源前的条件下,以足够的数量和纯度被生产出来。随后,在正确比例且无破坏性副反应的前提下,它们需被组装成核苷酸,并通过反复脱水缩合反应——在极不利于此类反应的水环境中——以精确序列连接数百至数千次。
现在让我们逐步审视:这三种关键成分是否可能在原始地球形成?若可能,此类事件的实际发生概率究竟有多大?
- 含氮碱的形成
含氮碱是具有复杂环状结构的复杂分子。这些分子从较简单的前生物化合物中自发,这是极不可能的,因为它需要特定的化学反应、特定的反应条件和催化剂才能形成环状结构。这些反应包括胺化反应,即在碳骨架上添加胺基(NH2),需要氨、醛或酮等含氮化合物,通常需要催化剂或高温的促进。除去氧原子的脱氧反应需要氢气或甲烷等还原剂。环的形成是形成含氮碱结构的关键,通常是在高温高压条件下进行的多步过程,通常由金属离子催化。最后,含氮碱的添加可能需要高能环境和特定的前体化合物才能完成。
据认为,地球早期的环境在温度、pH值和可利用的化合物方面差异很大。创造合成含氮碱基所需的精确条件是极具挑战性的。例如,形成这些碱基所需的高能条件可能无法持续存在或维持。即使在优化的实验室条件下,含氮碱基的产量也往往很低。这不禁让人怀疑,自然界是否能产生足够数量的这些碱基来支持RNA或其他核酸的形成。含氮碱基的合成途径涉及多个步骤和中间化合物。所有必要条件和化合物同时存在并达到正确比例的可能性值得怀疑。
含氮碱基的形成通常需要催化剂来驱动化学反应。在前生物世界中,这种催化剂在适当浓度和条件下的存在是不确定的。如果没有这些催化剂,反应速度将非常缓慢,无法产生重大影响。即使含氮碱能够自发形成,它们在前生物环境中的稳定性也是值得怀疑的。在紫外线辐射、水解和其他环境因素的作用下,这些分子很容易降解。这种不稳定性将阻碍它们的积累和随后用于形成RNA。
- 核糖的形成
福尔马林反应(Formose reaction)是一种在催化剂存在下甲醛发生聚合反应的过程,该反应可生成核糖。然而,该反应缺乏特异性,导致核糖的产量相对于其他糖类较低。此外,该反应需要特定条件,例如以氢氧化钙作为催化剂,而这些条件在前生物环境中可能并不普遍存在或稳定。为了使核糖在RNA的原始合成中发挥作用,它需要被选择性合成并稳定化。然而,福尔马尔反应并不利于核糖的选择性形成,由此产生的糖混合物使核糖在RNA合成中的利用变得复杂。因此,需要存在能够稳定核糖或从复杂混合物中选择核糖的机制。潜在的稳定剂,如硼酸盐矿物,已被提出,但其在前生物条件下的可用性和有效性尚不明确。
福尔马尔反应需要甲醛,其浓度必须足够高。在前生物条件下,甲醛的产生和稳定性是不可能的,因为甲醛容易聚合或与其他化合物反应。福尔马尔反应高效进行并产生核糖所需的特定环境条件(如最适pH值、温度、催化剂存在等)可能在早期地球上并不普遍或稳定。即使在受控实验室条件下,核糖的产率也较低,且反应会产生复杂的糖类混合物,这凸显了在前生物环境中分离核糖的挑战性。
核糖是一种戊糖,化学性质不稳定,容易快速降解,尤其是在地球早期的普遍条件下。之所以不稳定,是因为核糖在水溶液中很容易水解,并可通过马氏反应和焦糖化等过程降解。此外,研究还表明核糖的半衰期很短,尤其是在碱性条件下,因此不可能在地质时间尺度上大量积累。
- 磷酸基的形成
前生物条件下磷酸盐基团的形成面临着挑战,因为在早期地球上随时可用的磷酸盐来源相对匮乏。磷酸盐通常存在于磷灰石等矿物中,而磷灰石在水中的溶解度不高,因此磷酸盐很难在据认为发生了前生物化学反应的水环境中自由存在。在中性pH值条件下,磷酸盐矿物往往是化学惰性的。这种低反应性严重阻碍了磷酸盐与生命所需的有机分子的结合。
磷酸酯的形成对核苷酸的合成至关重要,需要大量的能量输入。在前生物条件下,克服这些障碍所需的能源和催化过程将是有限的。一些研究表明,高能条件(如雷击或火山活动产生的高能条件)可以促进含磷分子的形成。然而,这些情况需要特定的瞬时条件,而这些条件可能并不普遍。
聚磷酸盐是磷酸盐基团链,它的形成通常需要特定的条件,如高温或催化剂的存在,而在前生物环境中,这些条件可能并不容易获得。聚磷酸盐容易水解,分解成更简单的磷酸盐化合物。这些化合物在地球早期的波动条件下的稳定性值得怀疑。
虽然有些实验证明在模拟的前生物条件下形成了含磷酸盐的分子,但这些实验往往需要高度特定和受控的条件,可能无法真实反映早期地球的环境。此外,在前生物合成实验中,含磷酸盐分子的产量通常很低,这让人怀疑这些过程在前生物地球上以足以驱动生命起源的规模发生的效率和合理性。
- 功能RNA核苷酸的形成
即使克服了所有挑战,成功地创造了含氮碱基、核糖和磷酸基团,但另一个重大障碍依然存在:形成功能性RNA核苷酸。
RNA有多种类型:参与蛋白质合成的RNA(mRNA、rRNA、tRNA等)、参与转录后修饰的RNA(snRNA、snoRNA等)、调控RNA(aRNA、miRNA等)和寄生RNA。RNA分子中核苷酸的数量取决于其类型。例如
- mRNA和rRNA-数百到数千
- tRNA-70至90
- snRNA-100至300
- miRNA-20至25。
假设我们要估算形成概率的典型RNA分子长度为100个核苷酸。在这种情况下,RNA序列中的每个位置都可以由腺嘌呤、尿嘧啶、胞嘧啶或鸟嘌呤四种碱基中的一种占据。长度为100个核苷酸的可能序列总数为4100(=1.6x1060),形成功能RNA的概率为1/1.6x1060=6.2x10-61。这个极小的概率表明,即使存在原有的含氮碱基、核糖和磷酸基团,功能RNA也不可能自发形成。
iii.蛋白质的形成
蛋白质的形成包括氨基酸的合成、氨基酸聚合成肽以及这些肽折叠成功能性蛋白质。让我们来看看在前生物条件下这些过程中存在的问题和面临的挑战。
蛋白质由称为多肽链的氨基酸长链组成,这些氨基酸以高度特定的序列排列。单个蛋白质中的氨基酸数量从几十个到几千个不等。例如,小型蛋白质胰岛素含有约51个氨基酸,中型蛋白质肌红蛋白含有约153个氨基酸,大型蛋白质血红蛋白含有约574个氨基酸,而巨型蛋白质titin则含有约34350个氨基酸。由20种氨基酸随机组合形成长肽链几乎是不可能的。例如,小型蛋白质胰岛素通过随机过程形成多肽链的概率为1/2051=44x10-67≈0。
即使多肽链是以某种方式形成的,它们也必须折叠成特定的三维结构,才能成为功能性蛋白质。多肽链折叠成功能性蛋白质的过程涉及几个关键步骤,每个步骤都由各种化学作用驱动,并由细胞内的分子机器协助。
多肽链的某些段落(一级结构)折叠形成称为α螺旋和β折叠的二级结构。这些结构通过多肽链主链原子之间的氢键得以稳定。额外的二级结构,如转角和环,连接螺旋和片层,有助于蛋白质的整体折叠。二级结构进一步折叠成特定的三维形状,称为三级结构。这一过程由疏水作用驱动,其中非极性侧链远离水环境,促使多肽链折叠成紧凑的球形结构;氢键,形成于极性侧链与主链之间, 稳定折叠结构;离子键,通过带相反电荷的侧链之间的静电相互作用增强蛋白质的稳定性;以及二硫键,由半胱氨酸残基之间的共价键提供额外的结构稳定性。
对于某些具有多条多肽链(亚基)的蛋白质来说,这些折叠后的单位会聚集在一起形成四元结构。为了防止出现错误,伴侣蛋白通过防止错误折叠和聚集来协助折叠过程。它们帮助多肽链实现正确的构象。蛋白质可能会发生微小的构象变化和修正,以达到最稳定和功能性最强的构象。还可能发生磷酸化、糖基化或裂解等化学修饰,进一步稳定蛋白质或为其特定功能做好准备。
氨基酸之间肽键的形成需要大量能量。在前生物条件下,是否有持续而充足的能量来源来驱动这些反应值得怀疑。虽然有人提出了闪电、紫外线辐射和火山热等各种能源,但这些能源在持续促进肽键形成方面的效率和可靠性还值得商榷。地球早期的条件可能是恶劣和多变的,温度、pH值和环境变化都很极端。这些条件可能会破坏肽键形成的微妙过程和已形成肽的稳定性。
肽和氨基酸在水环境中会发生水解和降解。已形成的肽的长期稳定性令人担忧,因为它们的降解速度可能比形成速度更快。在前生物条件下缺乏保护机制意味着新形成的肽可能会被紫外线辐射和热波动等环境因素迅速分解。虽然粘土等矿物表面可以催化肽键的形成,但这些反应在自然条件下的效率、特异性和产量还没有得到充分证明。目前还不确定这些表面在生产生命所需的各种肽方面有多大作用。发生这些矿物催化反应的精确条件(如温度、pH值)必须严格控制,而早期地球上可能并不一直存在这样的条件。一些证明肽形成的实验是在高度受控的条件下进行的,但这些条件可能无法准确反映早期地球混乱多变的条件。
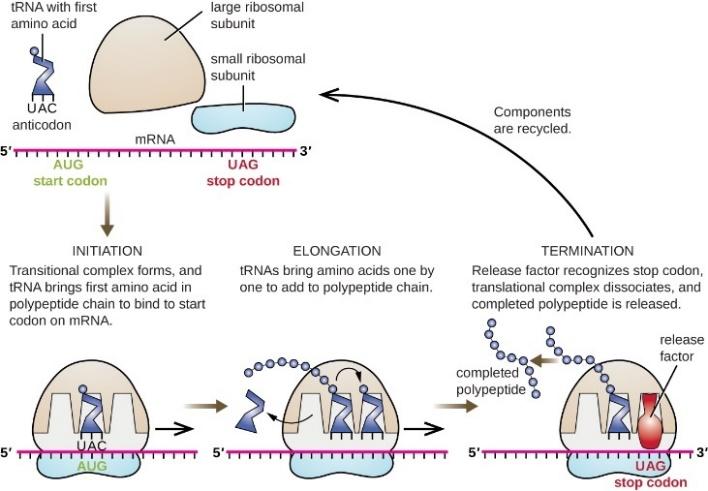
图3.2.蛋白质合成
RNA世界假说认为,RNA分子催化了肽的形成。然而,功能性RNA和肽的同时出现带来了一个“鸡生蛋、蛋生鸡”的问题,两者相互依存。没有RNA,就无法形成蛋白质。
蛋白质需要具有相同手性的氨基酸(L-氨基酸)。前生物合成通常会产生外消旋混合物,其中含有等量的左旋和右旋异构体。从统计学角度看,从这种混合物中自发形成同手性蛋白质是不可能的。
iv.DNA的形成
前生物条件下DNA的形成是一个复杂的推测过程,涉及几个关键步骤,包括核苷酸合成、多核苷酸链形成、碱基配对、双螺旋形成、DNA缩合、复制和酶辅助。
与RNA一样,DNA核苷酸也由三部分组成:含氮碱基(腺嘌呤、鸟嘌呤、胞嘧啶、胸腺嘧啶)、脱氧核糖和磷酸基团。自发形成DNA的难度与RNA相当。DNA的另一个难点是DNA双螺旋结构的形成。DNA的双螺旋结构依赖于腺嘌呤和胸腺嘧啶以及胞嘧啶和鸟嘌呤之间精确的碱基配对。在没有指导模板或机制的情况下,自发实现这种特异性是极其不可能的。要形成稳定的双螺旋,核苷酸必须按照特定的顺序排列,互补序列位于相反的链上。自发形成两个完全一致的互补序列的可能性极低。
DNA复制需要复杂的酶和蛋白质机制来确保准确性和保真度。参与DNA复制的关键酶包括螺旋酶、单链结合蛋白(SSB)、引物酶、DNA聚合酶、核糖核酸酶H(RNaseH)、DNA连接酶和拓扑异构酶。自发形成的双螺旋不包括这些基本成分,因此复制和纠错的可能性极低。如果没有纠错机制,任何自发形成的DNA都可能迅速积累错误,损害其稳定性和功能性。
参与DNA复制的典型酶的氨基酸总数在几百到几千之间。随机产生这些酶的概率几乎为零。例如,随机产生RNaseH的概率仅为20-155或2.2x10-202≈0。这种令人难以置信的小概率基本上超出了实际发生的范围,在自然界中永远不会发生。
即使DNA是以某种方式形成的,它也需要经过非常复杂的DNA缩合过程。DNA缩合过程将长长的线性DNA分子转化为高度紧凑和有序的结构,使其能够装入细胞核内。凝结过程对于DNA的有效储存、保护和调节以及细胞分裂过程中染色体的正常分离至关重要。这一过程包括形成核小体、30纳米纤维、环状结构域、高阶折叠和分裂相染色体。
如果DNA缠绕组蛋白,就会形成核小体。每个核小体由大约147个碱基对的DNA组成,包裹着组蛋白八聚体(H2A、H2B、H3和H4各两份)。由此产生的结构看起来就像串珠,核小体(珠子)由连接子DNA(线)连接。
在连接子组蛋白H1的作用下,核小体链进一步盘绕成更紧凑的30纳米纤维,组蛋白H1与核小体和连接子DNA结合。根据核糖体相互作用的不同,30nm纤维可采用螺线形或之字形构型。
30纳米纤维通过附着在细胞核内的蛋白质支架上形成环状结构域。支架或基质附着区(SARs/MARs)锚定这些环。这些环的长度通常为40-90千碱基对(kb),可进一步压实,并通过将遥远的调控元件带到基因附近,在基因调控中发挥作用。
环状结构域进一步折叠成更粗的纤维,即染色质纤维。这些纤维经过更多的卷曲和折叠,形成了更加紧凑的结构。
在细胞分裂过程中,尤其是在分裂后期,染色质会达到最高的凝结水平,从而形成可见的染色体。这涉及凝集素蛋白的作用,它们有助于超卷曲和压缩染色质。每条染色体都由两条相同的姐妹染色单体组成,并在中心粒处固定在一起,确保细胞分裂过程中的准确分离。
凝集程度会影响基因表达,紧密排列的异染色质转录不活跃,而松散排列的常染色质转录活跃。在有丝分裂和减数分裂过程中,适当的凝集对染色体的准确分离至关重要。
如上所述,DNA的形成和复制非常复杂,需要精确的生化协调和各种酶的参与。然而,进化论并没有明确解释这些机制是如何起源的,只是简单地说DNA是由RNA演化而来的,而没有解决关键的难题。要使这一说法成立,就必须解释RNA是如何形成的,DNA的双螺旋结构是如何出现的,以及重要的复制酶是如何起源的。没有这些答案,这一观点仍然是推测性的。考虑到这些因素,DNA的形成是有意设计的结果,而不是随机机遇。
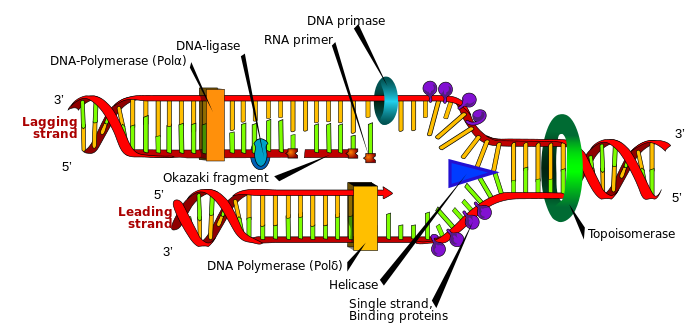
图3.3.DNA复制过程
v.细胞的形成
为了继续我们的讨论,让我们假设RNA、蛋白质和RNA是自发产生的。那么,生命的下一步就是细胞的形成。细胞主要有两种:原核细胞和真核细胞。原核细胞存在于细菌和古细菌等生物体内,比较简单,没有明确的细胞核。它们的遗传物质包含在单个环状DNA分子中,在细胞质中自由漂浮。原核细胞也缺乏膜结合的细胞器。真核细胞存在于植物、动物、真菌和原生动物中,结构更为复杂。它们包含一个由核膜包裹的明确的细胞核。真核细胞还拥有各种膜结合细胞器,如线粒体、内质网和高尔基体,这些细胞器发挥着细胞生存和正常运作所必需的特定功能。
科学家声称,原细胞是在自然选择、突变和环境适应的驱动下逐渐进化成原核细胞的。原细胞是现代细胞的假想前身,它的存在面临着一些重要的批评。其中一个主要问题是脂质双分子层的自发形成,这对于创造稳定的封闭环境至关重要。在早期地球上形成并持续维持这些双分子层所需的条件是高度推测性的。此外,在这些脂质结构中整合功能成分(如RNA或简单蛋白质)需要高度特异性的相互作用,如果没有某种引导机制,从统计学角度看是不可能的。此外,原细胞的复制和进化能力--生物体的一个关键特征--也缺乏足够的实验支持,这使人们对它们在生命起源中的作用产生了疑问。基于这些原因,地球上最早出现的细胞应该是原核细胞。
化石记录表明,原核细胞出现在35亿至38亿年前的地球上。所有细胞都被细胞膜所包围,而细胞形成的第一步就是细胞膜的形成。因此,让我们来研究一下细胞膜能否在前生物条件下自发形成。
- 细胞膜的形成
细胞膜并不简单,而是由脂质(磷脂、胆固醇和糖脂)、蛋白质和碳水化合物组成的复杂而动态的结构。磷脂形成基本的双层结构,胆固醇调节流动性,糖脂则有助于细胞识别。蛋白质(包括整体蛋白质和外周蛋白质)促进运输、信号传递和结构支持,而碳水化合物则在细胞识别和交流中发挥关键作用。这种成分使细胞膜能够发挥其基本功能,维持平衡并促进与环境的相互作用。
由于功能性膜结构所需的复杂性和特异性,在前生物条件下随机形成细胞膜面临着一些问题。
特定的两亲脂质分子,如磷脂,需要脂肪酸、甘油和磷酸基团的精确组合,而在前生物条件下,这些脂肪酸、甘油和磷酸基团不太可能以正确的比例自发形成和组装。磷酸基团的自发形成,如上一节所示,是不太可能的。虽然两亲分子可以自发形成双分子层,但要形成一个稳定的、半渗透性的、能够包裹和保护细胞环境的双分子层需要特定的条件。随机出现这些条件(包括适当浓度和类型的脂质)的可能性很小。
原核细胞(如细菌细胞)的典型大小为1微米。表面积为3x10-12m2,单个磷脂分子的大小约为5x10-19m2。因此,双分子层中磷脂的总数为1.2x107。要形成双分子层,大约1千万磷脂必须并排排列,形成一个封闭的腔室。这种情况极不可能随机发生,因为如果没有某种形式的引导或指导,双分子层是不会自然排列并形成一个封闭的腔室的。
地球早期的条件恶劣而多变,温度、pH值和辐射都达到了极致。在这样的环境中保持原始膜的完整性和稳定性是一项挑战,因为膜很容易被这些因素破坏。功能膜必须有选择性地允许必需的营养物质和分子通过,同时将有害物质拒之门外。这种选择性渗透性要求存在复杂的蛋白质和通道,它们不太可能通过随机过程形成并整合到膜中。
即使真的形成了原始膜,也不可能随机封装核苷酸、氨基酸和催化分子等必要的生物大分子。启动原始代谢过程所需的特定浓度和组合也不可能偶然出现。
功能膜的形成必须伴随着其他细胞机制(如运输蛋白和代谢酶)的同步发展,这使得膜形成的随机过程更加复杂。因此,在前生物地球上形成原核细胞是不可行的。
vi.真核细胞的形成
被广泛接受的真核细胞起源理论是内共生理论。内共生理论认为,真核细胞起源于原始原核细胞之间的共生关系。在这一过程中,祖先宿主细胞吞噬了某些原核细胞(动物细胞中的线粒体和植物细胞中的叶绿体),从而建立了互利关系,并最终形成了复杂的真核细胞。据说祖先宿主细胞是古细菌,但这一假说存在的问题是,在古细菌中从未观察到吞噬原核细胞的内吞作用,而且古细菌的细胞膜是由醚键组成的,而真核细胞的细胞膜是由酯键组成的。
这一理论要求预先存在原核细胞和线粒体或叶绿体。然而,线粒体和叶绿体的起源并没有很好的记载。线粒体是一种复杂的细胞器,具有独特的结构,反映了其作为细胞动力室的作用,通过氧化磷酸化产生ATP。线粒体由几个不同的部分组成:外膜、膜间隙、内膜和基质,其中包括酶、DNA、核糖体和代谢物。外膜与细胞膜一样,含有磷脂双分子层,其中混有磷脂和蛋白质。这种复杂的结构不可能通过随机过程自发产生,因为细胞膜、DNA和蛋白质不可能自发形成。线粒体有自己的DNA,不同于核DNA,但它们必须与核基因组协调才能正常工作。将线粒体DNA整合到宿主细胞的调控和代谢网络中是一项重大挑战。
真核细胞的细胞核由双层核膜、核仁和染色体组成,其中包含细胞的遗传物质,包括DNA、RNA和相关蛋白质。真核细胞中细胞核的起源更难解释。让我们从最简单的方面开始讨论:核膜。真核细胞中核膜的起源是科学界争论的焦点。人们提出了几种假说,包括膜内陷(向内折叠)假说、病毒起源假说和基因转移假说,来解释这种复杂结构是如何产生的。
膜内陷假说认为,核膜起源于祖先原核细胞的细胞膜内陷。然而,这一假说无法解释细胞膜与核膜之间的区别。细胞膜由单层磷脂双分子层组成,而核膜由两层磷脂双分子层组成--内膜和外膜。此外,核膜还含有细胞膜所没有的核孔复合体。此外,细胞膜和核膜中的蛋白质组成也不同。
病毒起源假说认为,感染原始细胞的病毒可能提供了遗传物质或结构成分,最终导致核膜的形成。病毒和宿主细胞膜之间的相互作用可能在DNA周围形成了一个保护结构。虽然已知病毒会影响宿主细胞的结构,但将病毒与核膜起源联系起来的具体证据却很有限。
基因转移假说认为,不同原核生物之间基因的混合和转移可能会产生一个庞大而复杂的基因组,需要一个保护区。核膜的进化就是为了保护和调节这种复杂的遗传物质。这一假说面临许多问题,因为它缺乏直接证据,无法解释如何仅通过基因的转移和整合就产生了如此复杂和有组织的双层膜和核孔复合体结构,也未能提供明确的途径说明转移的基因如何以一种导致核膜发展的方式进行整合和表达。
核小体和染色体的结构要比核膜复杂得多,因此很难想象它们是由随机事件产生的。此外,要了解这些成分是如何被包裹在核膜中的,也是一项挑战。核小体和染色体包含生物体的遗传信息,包括形成RNA、蛋白质、DNA、细胞器以及生物体组织和器官的蓝图。在真核细胞阶段,甚至在生命形成之前,这些构建生命的蓝图就已经被预测并存在于细胞核中,这一事实是进化论无法充分解释的。相反,这恰恰是智慧设计生命的明证。
总之,智能设计可以自然地解释真核细胞的起源,而进化论对真核细胞的起源缺乏明确的解释。
vii.细胞器定位
细胞由各种细胞器组成,包括细胞核、线粒体、内质网、高尔基体、溶酶体和其他细胞器,它们共同维持细胞功能和平衡。细胞器定位是一个高度调节的动态过程,可确保细胞器在细胞内的最佳位置,以维持高效的细胞功能。正确的定位对细胞健康至关重要,并在适应不断变化的细胞和环境条件方面发挥着关键作用。人们可能会问,这些细胞器无法自我思考,它们是如何找到最佳位置的?
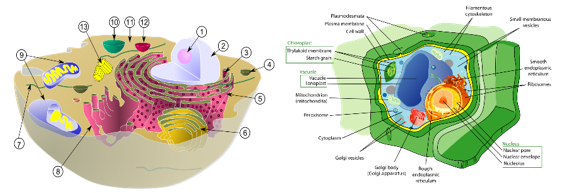
图3.4.动物细胞和植物细胞的结构
对细胞器定位过程的详细研究揭示了一个高度精确和错综复杂的机制,它不能归因于随机的机会。这一过程涉及细胞骨架、运动蛋白、膜贩运、锚定蛋白、支架、动态调整和细胞器间通信等复杂的相互作用。
细胞骨架在细胞器定位中起着至关重要的作用。它提供结构支持,促进运动,并确保细胞器的正确定位。细胞骨架主要由三类丝组成:微管丝、肌动蛋白丝和中间丝。
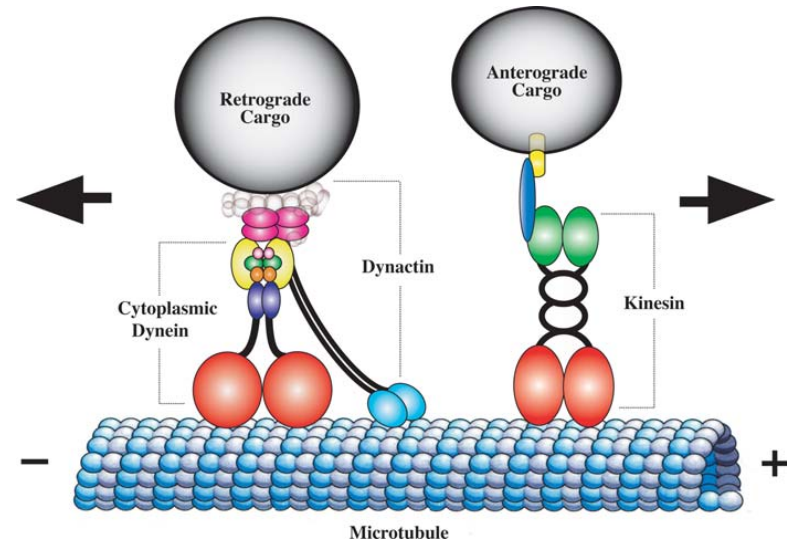
图3.5.微管和马达蛋白示意图
微管是由微管蛋白组成的中空长管。它们形成一个网络,从微管组织中心(中心体)延伸到细胞外围。微管是运动蛋白(如驱动蛋白和动力蛋白)的轨道。驱动蛋白将细胞器移向微管的正端,通常移向细胞外围,而动力蛋白则将细胞器移向负端,通常移向细胞中心。微管有助于定位细胞器,如高尔基体(通常位于中心体附近)和线粒体,线粒体分布在整个细胞中,但可以沿着微管运送到能量需求高的区域。
肌动蛋白丝又称微丝,是由肌动蛋白蛋白质组成的纤细柔韧的纤维。它们集中在质膜下方,并在整个细胞质中形成密集的网络。肌动蛋白丝促进细胞质流动,这一过程有助于细胞器和营养物质在整个细胞内的分布。肌球蛋白运动蛋白与肌动蛋白丝相互作用,沿着肌动蛋白网络运输囊泡、内体和其他小型细胞器。肌动蛋白丝有助于维持细胞形状并参与细胞运动,从而间接影响细胞器的定位。
中间丝是由各种蛋白质(如角蛋白、波形蛋白和层粘连蛋白)组成的绳状纤维,取决于细胞类型。它们提供机械强度和结构支持。中间丝通过将细胞核等细胞器固定在细胞质内的位置,帮助它们稳定位置。它们能保持细胞骨架的整体完整性,确保微管和肌动蛋白丝等其他成分能在细胞器定位过程中有效发挥作用。
不同类型的细胞骨架丝通常相互配合,以准确定位细胞器。例如,微管和肌动蛋白丝相互协调,确保囊泡和细胞器的正确分布和移动。细胞骨架是高度动态的,不断重塑以适应细胞的需要。这种灵活性使细胞器能够根据细胞信号或环境变化迅速重新定位。
膜运输是蛋白质、脂质和其他分子在细胞内的运输过程,确保细胞成分到达正确的目的地。这包括囊泡从供体膜上萌发、通过细胞质运输以及与目标膜融合。参与膜运输的主要细胞器包括内质网、高尔基体以及各种类型的囊泡,如内体和溶酶体。这一过程对于维持细胞组织、促进细胞器之间的交流以及使细胞对内部和外部信号做出有效反应至关重要。
信号通路引导细胞器在细胞内的运动和定位。这些途径涉及化学信号的传递,这些信号提供空间线索,确保细胞器被引导到适当的位置。细胞器表面和细胞质内的受体与信号分子相互作用,促进这一过程。例如,Rab蛋白等小GTP酶是通过与特定效应蛋白相互作用来控制囊泡贩运和细胞器定位的关键调节因子。这些信号通路确保了细胞过程的协调,以及细胞器的动态定位,以应对不断变化的细胞需求和环境条件。
锚蛋白和支架在细胞定位中发挥着重要作用,它们确保细胞器在细胞内精确定位。锚定蛋白将细胞器连接到细胞质内的特定位置,使其稳定并防止移位。例如,线粒体可以通过特定的锚定机制拴在内质网上,从而促进有效的能量转移和代谢协调。支架蛋白通过形成复合物,将细胞器固定到位,从而提供结构支持,维持细胞的整体组织。这些蛋白质创建了一个动态框架,使细胞器能够正确排列,确保细胞功能有效、高效地发挥。
细胞定位的动态调整是指细胞内细胞器定位的持续和反应性变化。这些调整对于维持细胞功能和适应性至关重要。在细胞周期的不同阶段,如有丝分裂,细胞核和线粒体等细胞器会重新定位,以确保细胞正常分裂。此外,为了应对营养供应或压力条件等环境刺激,细胞器还可以重新定位到最需要其功能的区域。细胞骨架和运动蛋白为这种动态重新定位提供了便利,使细胞能够保持平衡并有效地应对不断变化的内部和外部条件。
细胞器之间的交流确保了细胞功能的协调和效率。这种交流通过直接接触点和囊泡运输进行。接触点,如线粒体和内质网之间的线粒体相关膜(MAM),可促进脂质、钙和其他分子的转移,确保细胞器之间的同步活动。囊泡运输涉及囊泡的脱落和融合,囊泡在细胞器之间携带蛋白质和脂质,维持细胞器的功能整合。有效的细胞器间通讯对于新陈代谢、信号传递和应激反应等过程至关重要,有助于细胞的整体平衡。
如上所述,细胞器定位所涉及的机制是高度有序和复杂的。由于以下原因,通过随机突变和自然选择逐步演化出这种错综复杂的协调系统的可能性极小。
没有直接证据表明细胞器定位机制进化的中间阶段。化石记录和分子研究并没有捕捉到能够说明这些复杂系统逐步进化的过渡形态。细胞器定位及其在细胞内协调的复杂性对进化论的解释提出了挑战,因为细胞组织呈现出“不可还原的复杂性”,去除任何部分都会使系统失去功能。进化理论通过渐变来解释复杂性,但细胞结构及其精确定位没有可行的中间阶段。
细胞器的定位取决于与细胞骨架、运动蛋白、信号通路和其他细胞成分之间错综复杂的相互作用。这种相互依存的关系提出了这样的问题:这些系统,是如何以循序渐进的方式共同进化的?要解释细胞器和负责其定位的系统是如何同时进化的,而不是先有一个系统完全发挥作用,是很有挑战性的。
人们对驱动蛋白、动力蛋白和肌球蛋白等运动蛋白以及微管和肌动蛋白丝等细胞骨架元素的起源和进化并不完全了解。这些蛋白质和结构一定进化出了高度特异的功能和相互作用,这很难仅仅通过渐进的变化来解释。控制细胞器定位的复杂调控网络的进化带来了重大挑战。这些网络必须精确地协调众多基因的表达和活动,而它们通过随机突变的渐进进化很难解释。
参与细胞器定位的许多成分是相互依存的,这意味着它们必须有效地共同发挥作用,才能提供任何选择性优势。多个相互影响的部分同时进化是有问题的,因为部分系统不会带来足够的益处而受到自然选择的青睐。
细胞器的定位和维持过程是能量密集型的。目前还不清楚,如果没有高效的能量生产和资源管理机制,早期细胞如何能够承受与这些复杂系统相关的代谢成本。
viii.细胞分化
细胞分化是指非特化细胞发展成为具有独特结构和功能的特化细胞的过程。这一过程对于组织、器官以及最终的多细胞生物体的发育、生长和功能至关重要。分化通常从干细胞开始,干细胞是未分化的细胞,能够产生各种类型的细胞。干细胞具有全能性,几乎可以分化成任何细胞类型。在发育过程中,这些细胞接收信号,引导它们成为特定类型的细胞。随着干细胞的分化,它们成为多能祖细胞,致力于产生有限范围的细胞类型。祖细胞进一步分化为完全特化的细胞。细胞分化是一个受基因表达调控、信号转导途径、表观遗传修饰、形态发生梯度以及与其他细胞和细胞外基质相互作用驱动的高度调控的动态过程。
生物体内的所有细胞都含有相同的DNA,但不同类型的细胞表达不同的基因子集。这种选择性基因表达推动了细胞分化。被称为转录因子的蛋白质与特定的DNA序列结合,调节目标基因的转录。这些因子可以激活或抑制基因表达,从而产生特定细胞类型所需的蛋白质。
细胞接收来自环境的信号,如生长因子、激素和细胞因子。这些信号与细胞表面受体结合,启动信号转导途径。信号转导途径涉及一连串的细胞内事件,通常包括蛋白质的磷酸化,最终导致基因表达的变化。
表观遗传修饰包括DNA甲基化和组蛋白修饰。DNA甲基化通过向DNA(通常是CpG岛)添加甲基来抑制基因表达。甲基化模式具有遗传性,可通过抑制特定细胞类型不需要的基因来锁定细胞的特性。组蛋白是缠绕DNA的蛋白质,可以进行化学修饰(如乙酰化、甲基化)。这些修饰可改变染色质结构,使DNA可用于转录。
形态发生因子是一种信号分子,可在组织中扩散并形成浓度梯度。细胞对不同浓度的形态发生因子做出反应,激活不同的发育途径,导致不同的细胞命运。在胚胎发育过程中,形态发生梯度对模式形成至关重要,它决定了分化细胞的空间排列。
细胞之间的直接接触可诱导分化。一个细胞上的膜结合蛋白与邻近细胞上的受体蛋白相互作用,传递信号。细胞分泌的信号分子会影响附近的细胞,从而影响它们的分化。
细胞外基质(ECM)由蛋白质和多糖组成,为细胞提供结构支持和生化信号。整合素和其他粘附分子介导细胞与ECM的粘附,影响细胞的形状、迁移和分化。
正反馈和负反馈机制控制着分化进程。正反馈表明,分化的细胞可以产生强化其特征的信号,确保细胞类型的稳定。负反馈机制限制分化信号,防止分化过度,维持未分化细胞池。
如前所述,细胞分化涉及一系列高度复杂和协调的事件,包括精确的基因调控、信号转导和表观遗传修饰。这种复杂性很难仅仅通过渐进的随机突变和自然选择来解释。这一过程需要众多细胞系统的整合,如转录因子、信号通路和细胞骨架。这些相互依存系统的同时进化对进化理论提出了重大挑战。此外,多能干细胞的起源也无法用进化机制来解释。
表观遗传修饰(如DNA甲基化和组蛋白修饰)在分化过程中起着至关重要的作用。进化理论无法很好地解释这些复杂机制的起源,因为它们需要高度的精确性和协调性。表观遗传标记的遗传性又增加了一层复杂性。这些标记的建立、维持和遗传机制错综复杂,需要详细的解释。
形态发生梯度的建立和解读对于发育过程中的模式形成至关重要。精确的浓度梯度和细胞准确解读这些信号的能力表明,这是智能设计,而不是随机突变。位置信息的概念,即细胞确定自己的位置并进行相应的分化,需要一个复杂的通信系统。目前还不清楚这种系统的进化起源。
分化过程中控制基因表达的转录因子调控网络非常复杂。由于需要多个基因协调变化,这些网络的渐进进化缺乏经验支持。关键转录因子的突变会产生广泛的有害影响,因此很难设想有益的突变是如何逐渐累积形成功能性调控网络的。
ix.组织和器官的形成
组织的形成(组织发生)是胚胎发育过程中分化细胞组织成特定组织的过程。
在这个过程中,干细胞分化成各种类型的细胞,如肌肉细胞、神经细胞和上皮细胞,每种细胞都具有不同的功能。细胞分化后,开始排列成复杂的结构,形成人体的基本组织。这些组织包括上皮组织、结缔组织、肌肉组织和神经组织,每种组织都对器官的整体结构和功能做出了贡献。
细胞通信和信号通路在引导细胞到达正确位置并确保它们适当互动方面发挥着至关重要的作用。组织发生受到严格调控,因为细胞组织错误会导致发育异常或疾病。在整个过程中,细胞相互粘附,迁移到特定区域,并发生形态变化,形成功能性组织结构。组织发生过程完成后,就形成了能够执行特殊功能的发育完全的组织。这一过程是器官正常发育和人体整体组织的基础。
器官的形成(器官发生)是在组织发生之后进行的,在组织发生过程中,组织被组织成功能单元。在器官形成过程中,三个胚层--外胚层、中胚层和内胚层--相互作用并进一步分化,形成特定的器官。外胚层主要形成大脑和脊髓等器官,而中胚层则形成心脏、肾脏和骨骼肌。内胚层形成肺和肝脏等内部结构。
器官发生涉及复杂的信号通路和基因调控,以确保器官在正确的位置发育并具有适当的功能。在器官发生过程中,细胞会根据需要迁移、增殖和凋亡,以形成发育中的器官。Notch信号通路在决定细胞命运和维持细胞增殖与分化之间的平衡方面尤为重要。Wnt信号通路有助于器官的模式化和形态发生,确保组织以正确的位置和比例发育。这些信号的中断会导致先天性缺陷或器官发育异常。这一过程对于建立人体的整体解剖学和生理学至关重要。
随着器官的发育,多种组织类型会整合在一起并共同发挥作用。例如,像心脏这样的器官由肌肉组织、结缔组织和神经组织组成,所有这些组织对其功能都至关重要。这些器官的发育由复杂的信号通路引导,以确保细胞迁移到正确的位置、适当分化并形成正确的结构。
解释组织和器官形成的进化理论面临重大挑战。组织和器官过于复杂,无法用循序渐进的进化过程来解释。许多组织和器官表现出“不可还原的复杂性”,这意味着它们由多个相互依存的部分组成,缺少任何一个部分都无法发挥作用。这种复杂的结构不可能是逐步进化而来的,因为它们在中间阶段就会失去功能。
进化论认为,组织和器官等新结构是通过逐步改变现有结构而产生的。然而,这并不能充分解释没有明显前体的全新结构的起源。例如,大脑或免疫系统等复杂器官的发展就很难通过微小、渐进的变化来解释。
构建和组织组织和器官所需的遗传信息非常庞大,而且具有高度特异性,不可能通过随机突变产生如此详细的信息。
表观遗传因素在不改变DNA序列的情况下影响基因表达,在组织和器官的发育过程中发挥着重要作用。进化论主要强调基因突变,并不能完全解释表观遗传调控所带来的复杂性。该理论也无法解释复杂的生物系统(由多个相互作用的组织和器官组成)是如何独立进化,然后,整合成为一个统一的有机体。
x.多细胞生物的形成
一旦单个个器官形成,它们必须整合成一个具有凝聚力和功能性的有机体。这种整合是通过体内器官的空间组织来实现的,每个器官都占据一个特定的位置,使其能够与其他器官和系统相互作用。例如,包括心脏和血管在内的循环系统必须与呼吸系统和消化系统等其他系统妥善连接,才能维持生命。
在这一过程中,组织和器官内的细胞不断特化并适应各自的角色,这一过程被称为功能分化。这确保了生物体的每个部分都能有效地履行其指定的功能。不同器官和系统之间的协调和相互作用对于维持多细胞生物体的整体健康和功能,使其能够生存、生长和繁殖至关重要。从进化角度解释由器官形成多细胞生物体的过程,需要解决几个关键的难题和复杂性:
由器官形成多细胞生物体需要各个系统之间高度的整合与协调。导致多个器官系统同时发育和无缝运作的进化过程难以解释。
多细胞生物体内的器官和系统是高度相互依存的,这意味着一个系统的功能往往取决于其他系统的正常运作。进化论必须解释不同器官和系统的同时发展,每个器官和系统都有特定的功能和相互依存关系,并解释这些复杂系统是如何以协调、逐步的方式进化的。只有部分系统得到发展的中间形态不会提供足够的优势,从而受到自然选择的青睐。
化石记录中缺乏清晰的过渡形态,无法说明简单的多细胞生物逐渐进化为具有完整器官的复杂生物的过程。这一空白使得人们难以追溯导致这种复杂结构发展的进化途径。
器官形成和整合所需的基因表达和发育途径的精确协调是一项重大挑战。这些过程中的微小失误都可能导致发育障碍,这就提出了这样一个问题:这种微妙的系统是如何逐步进化的?
复杂的多细胞生物体的发展需要强有力的机制来处理错误和变异。进化论的解释必须说明这些错误处理系统是如何进化的,以及它们是如何确保器官形成和功能的稳定性和保真性的。
b.进化论能解释生命的起源吗?
在上一节中,我们讨论了生命的起源,追溯了从氨基酸、核糖核酸、蛋白质、脱氧核糖核酸、原核细胞、真核细胞、组织和器官的形成到最终形成多细胞生物体的过程。不可否认的是,这些过程都是朝着一个单一的,即形成生物体。
这就提出了一个重要问题:进化是通过不定向的随机过程进行的,它能充分解释这些复杂的发展和生命的起源吗?针对这一问题,进化论科学家提出了各种理论。进化论的主要理论包括自然选择、突变、基因漂移和水平基因转移。让我们来简要了解一下这些理论。
自然选择是指具有优势性状的个体能够更成功地存活和繁殖,从而使这些性状在群体中世代相传。自然选择作用于生物体内现有的变异。因此,生命的起源及其基本组成元素(氨基酸、核糖核酸、蛋白质、脱氧核糖核酸)和结构(细胞、组织、器官和多细胞生物体)的形成需要自然选择以外的解释,因为这些过程缺乏选择发挥作用的必要先决条件(复制和功能)。
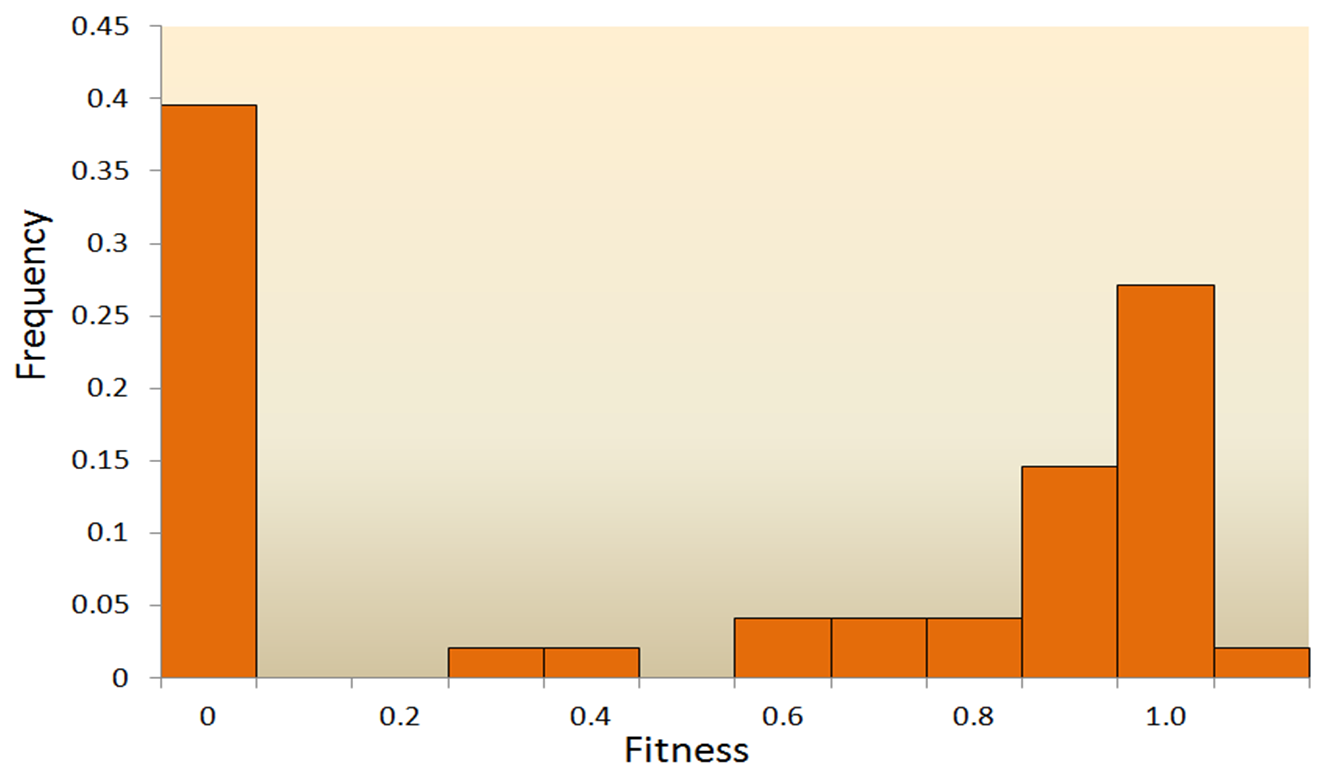
图3.6.健身效果的分布
突变是生物DNA中的随机变化,可带来遗传变异,有时会导致新的性状或适应性。突变面临着挑战,因为大多数突变都是有害或中性的,而不是有益的,这使得有利的突变不太可能频繁发生,以至于无法推动重大的进化变化。例如,一项关于水泡性口炎病毒随机突变的适应性效应分布(DFE)的研究就说明了这一问题。在所有突变中,39.6%为致命突变,31.2%为非致命有害突变,27.1%为中性突变。
如果核苷酸被插入或删除(导致移帧突变),或者停止密码子因突变而产生或消失,就会产生无功能的蛋白质。这就是为什么考虑到生物体蛋白质中的氨基酸数量众多(例如,人类蛋白质中的氨基酸数量从20个到33000个不等),不可能通过这种随机突变发生宏观进化的主要原因(详情参见本章“d”节)。此外,随机变异也无法解释生命最初从非生物物质中产生的原因。
遗传漂变依赖于等位基因频率的随机变化,这可能不足以解释在生物体内观察到的适应复杂性。遗传漂变在小种群中更为明显,因此在大多数进化都发生在大种群中时,它的影响就不那么重要了。此外,遗传漂变缺乏解释高度组织结构和系统发展所需的定向力。此外,遗传漂变不能产生新的信息或功能,因此无法解释新性状的出现或复杂生物特征的起源。
水平基因转移(HGT)是指遗传物质在不相关的生物体之间的转移,而不是通过遗传,从而导致基因变异。在解释多细胞生物的复杂性状时,HGT面临着一些问题,因为HGT的作用主要局限于原核生物,对高等生物的影响较小。将外来基因整合到宿主基因组中往往需要精确的调控机制,而这些机制不太可能同时进化。此外,HGT还会带来遗传不稳定性,可能导致有害突变。通过HGT获取基因的随机性也使人们对其产生协调和功能性适应的能力产生疑问。HGT无法解释新基因的起源,只能解释现有基因的转移,无法解释新性状的出现。
下表总结了进化理论对生物发生和遗传过程的适用性。
进化理论 | 能解释生物起源吗? | 能否解释RNA、蛋白质和DNA的形成? | 基因适应,而非进化?* |
自然选择 | 没有 | 没有 | 是 |
突变 | 没有 | 没有 | 是 |
遗传漂移 | 没有 | 没有 | 是 |
水平基因转移 | 没有 | 没有 | 不适用 |
表3.2.进化理论:生物发生和遗传学的适用性(*:遗传适应见下一节)
如表所示,主要的进化理论无法解释地球生命的起源,也无法解释核糖核酸、蛋白质和脱氧核糖核酸等基本生物成分的形成机制。这表明,应用于细胞、组织、器官和现有生命形式的进化模式并不能真正解释生命本身的起源或进化。这些理论并没有解决生命从非生物物质中产生的问题,而只是描述了一旦基本组成元素--RNA、蛋白质和DNA--已经到位,生命是如何发展的,就像详细描述汽车的组装过程或建筑物的建造过程,却没有解释原材料和部件是如何产生的。
应用于生物体的进化理论主要描述了使生物体能够适应不断变化的环境的遗传和生化过程。然而,这些适应性和行为并不是进化新创造出来的,而是已经编码在它们的遗传信息中的。鉴于这一局限性,进化论更准确的说法应该是“遗传适应论”(见下一节),因为它们主要解决的是生物如何通过已有的遗传机制来适应环境压力的问题。
尽管存在这些关键的局限性,进化论仍被过度推广,造成了广泛的误解。现在,许多人误以为进化论可以解释从非生命物质到生命有机体的转变以及复杂生命形式的发展。
要建造一座大楼,我们需要蓝图、建筑材料和坚实的地基。进化论就好比试图在没有蓝图(方向性)、建筑材料(RNA、蛋白质、DNA)和地基(生命的最初起源)的情况下建造一座大楼。没有这些,就无法建造大楼。
正如我们承认建筑蓝图是由建筑师设计的一样,我们也应该承认,所有生物都是由神圣的造物主上帝设计和创造的。
c.达尔文的理论:进化论还是遗传适应论?
进化论大致可分为两种类型:微进化与宏进化。微进化指的是物种在较短时间内发生的微小变化。这些变化可在较短时间内观察到,且通常涉及对环境的适应。宏进化则指在漫长的地质时期内发生的重大变化,最终导致新物种的形成及更广泛的分类群的出现。
进化生物学家认为,宏观进化的主要机制是微进化变化在时间上的积累。人们普遍认同微进化的证据存在,但缺乏宏观进化的确凿证据。若达尔文主义被称为进化论,必须提供宏观进化的证据。宏观进化的最有力证据是过渡物种的存在。达尔文在《物种起源》第六章(理论的困难)中写道:“如果物种是通过微小渐进的变化从其他物种演化而来的,为什么我们没有在各地看到无数的过渡形式?”这种过渡物种证据的缺失常被称为“达尔文的困境”。
被标记为“过渡”的化石可能仅仅是物种内的变异,或是完全无关的形态。这种模糊性使得难以确切识别真正的过渡形式。例如,提克塔利克(Tiktaalik)被广泛认为是过渡化石,并被视为脊椎动物进化研究中最重大的发现之一。然而,Niedzwiedzki等人发表在《自然》杂志上的论文揭示了保存完好的四足动物足迹,其年代比提克塔利克早约1800万年。这些足迹表明,完全发育的四足动物早在之前认为的时间之前就已经在陆地上行走。由于提克塔利克的年代约为3.75亿年前,这些更古老的四足动物足迹挑战了其作为鱼类与四足动物之间直接过渡形式的地位。
如果没有过渡物种的有力证据,达尔文的理论被误命名,应称为“遗传适应理论”而非“进化理论”。原因与米兰科维奇周期有关,该周期影响气候模式,并在塑造遗传适应过程中发挥了作用。
- 米兰科维奇周期
地球的偏心率在10万年周期内从近圆形逐渐变为椭圆形。偏心率的变化影响气候模式,影响冰河期和间冰期的时机。
地球的轴倾角(倾斜度)在4.1万年周期内变化范围为22.1度至24.5度。这种倾斜影响赤道与两极之间太阳辐射的分布,影响季节强度,并在长期气候模式和冰河时代动态中发挥关键作用。
地球自转轴的进动是指自转轴方向在26000年周期内逐渐改变。这种摆动导致季节的时序相对于地球在轨道上的位置发生偏移。这一机制改变了季节的强度和时序,影响地球整体气候系统。
地心距、地轴倾角和自转轴进动的综合影响被统称为米兰科维奇循环。这些循环导致全球气候的长期变化。撒哈拉沙漠是气候变化的典型例子。在太阳辐射增强的时期,撒哈拉地区降雨量增加,转变为一片郁郁葱葱、湖泊河流纵横的绿洲。相反,太阳辐射减少导致干旱条件,使该地区演变为今日所见的广袤沙漠。
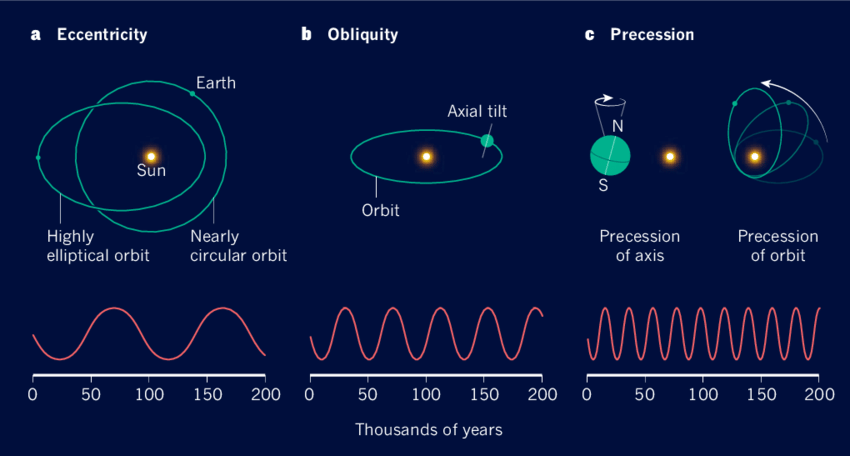
图3.7.米兰科维奇周期的组成部分
当此类变化发生时,地球上所有生物通过遗传适应调整自身以适应环境变化。这一非凡机制以DNA形式编码,使生物能够在漫长时期内生存而不至于灭绝。虽然进化论者传统上将这种适应性称为“进化”,但这种分类具有误导性;从科学角度而言,应更准确地将其描述为“遗传适应”。以下是一些能够支持“遗传适应理论”概念的例子。
- 遗传适应紫外线辐射
若人类皮肤因气候变化暴露于强紫外线辐射,一种涉及多种蛋白质和激素的复杂机制将通过激活特定基因,触发黑色素生成增加。
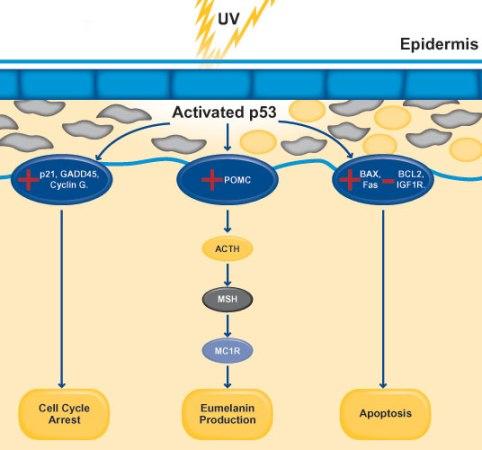
图3.8.黑色素生成机制
紫外线辐射会导致皮肤细胞的DNA损伤。这种损伤会激活p53蛋白,它是细胞应对压力和损伤的关键调节因子。激活的p53蛋白作为转录因子,促进参与紫外线损伤保护反应的多种基因的表达。p53刺激促黑素原(POMC)基因的表达。POMC是一种前体多肽,可被裂解为具有不同功能的多个小肽。POMC被加工成多种肽,包括促肾上腺皮质激素(ACTH)和黑素细胞刺激素(MSH)。
MSH与黑素细胞表面黑素皮质素1受体(MC1R)结合。黑素细胞是产生黑色素的细胞。MSH与MC1R结合后激活受体,触发黑色素细胞内的信号传导级联反应。MC1R的激活导致参与黑色素合成相关基因的上调。黑色素细胞增加黑色素的合成,这种色素可吸收并散射紫外线,从而保护皮肤细胞的DNA免受进一步的紫外线损伤。
黑色素被包装入黑色素小体,随后运输至皮肤外层的主要细胞类型:角质形成细胞。黑色素在角质形成细胞核周围形成保护性帽,有效屏蔽DNA免受紫外线辐射。
这是基因在相对较短时间内对环境变化作出适应性调整的典型案例。
- 基因适应北极环境
因纽特人通过遗传适应在严酷的北极环境中得以繁衍生息。关键适应包括脂肪酸去饱和酶(FADS)基因群的变异,这增强了他们从传统高脂肪的海洋哺乳动物饮食中代谢ω-3和ω-6脂肪酸的能力。此外,肉碱棕榈酰转移酶1A(CPT1A)基因的遗传变化提高了脂肪的能量转化效率,这对维持体温至关重要。这些适应性特征降低了高脂肪饮食带来的心血管疾病风险。此外,调控棕色脂肪活性的基因适应性增强了产热能力,帮助因纽特人在极寒环境中产生热量并维持体温。这些遗传适应性共同支持了他们在寒冷气候中的生存。这些变化似乎可追溯至至少2万年前,当时因纽特人的祖先生活在俄罗斯与阿拉斯加之间的白令海峡附近。这是遗传适应环境变化的又一例证。

图3.9.基因适应寒冷环境的因纽特人
- 棕熊到北极熊的遗传适应
棕熊向北极熊的演化是环境压力驱动遗传适应的典型案例。约40万年前,一群棕熊在北极地区被隔离,面临截然不同的生存挑战。适应严酷冰冻环境的遗传变异在自然选择中逐渐积累。
关键适应包括与脂肪代谢相关的基因变化,例如载脂蛋白B(APOB)基因,该基因改善了熊处理海豹高脂肪饮食的能力,而海豹是它们的主要食物来源。基因如内皮素受体B型(EDNRB)和黑色素瘤缺失1(AIM1)的适应性变化也导致了白色皮毛的形成,为熊在雪地和冰面上提供了伪装。此外,影响熊的骨骼结构和肢体形态的遗传变化增强了它们的游泳能力,这对在北极水域狩猎至关重要。
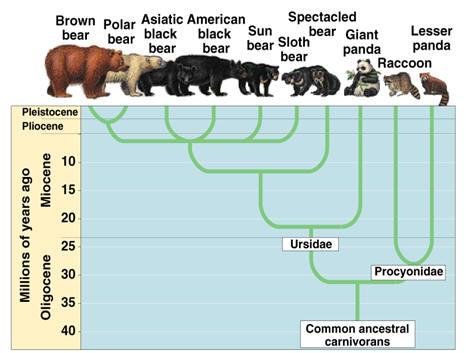
图3.10.棕熊和北极熊
这些遗传适应使北极熊能够高效利用北极资源,在极端寒冷环境中生存,并与棕熊祖先形成明显差异。值得注意的是,尽管经历了40万年的遗传变化,它们仍属于熊科,并未演化为不同物种。
- 雀类喙部形态的遗传适应
达尔文雀喙的大小和形状变化是遗传适应对环境压力的经典案例。在加拉帕戈斯群岛,雀类通过改变喙的形态以适应不同的食物来源。在干旱时期,当坚硬的种子成为主要食物来源时,喙较大较强的雀类更具选择优势并能繁殖后代。相反,当环境转向有利于较软食物时,喙较小、更灵活的雀类具有选择优势。这些适应是特定基因变化的结果,例如影响喙形状的无喙状同源盒1(ALX1)基因,以及影响喙大小的高移动性组AT钩2(HMGA2)基因。
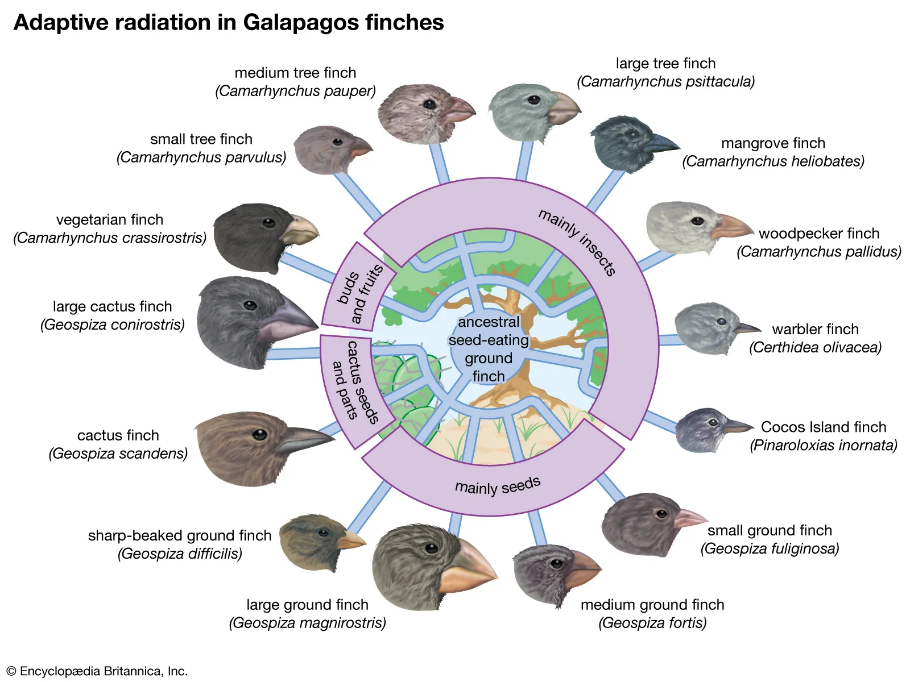
图3.11.加拉帕戈斯雀的鸟嘴
环境变化作用于这些遗传变异,导致适应不同生态位的多样化喙部形态。经过代际传递,这些遗传适应使雀类能够高效利用可用资源,从而展示了遗传变异如何在环境挑战下驱动喙部形态与大小的多样化。雀类在加拉帕戈斯群岛生活了约200万年。尽管经历了漫长时期,它们仍保持雀类形态,未演化为不同物种(即无宏观进化)。
综上所述,达尔文的“进化论”应称为“遗传适应理论”,因缺乏宏观进化的确凿证据。微进化指的是在一定时间内,种群内等位基因频率的小规模变化,而遗传适应则特指那些能增强生物在环境中生存和繁殖能力的遗传变化。因此,在讨论赋予生物生存优势的变化时,使用“遗传适应”这一术语更为恰当和准确。
d.我们是从人猿进化而来的吗?
人类学家认为,人类进化始于约2040万年前的类人猿(Hominoidea)。类人猿随后分化为人科(Hominidae)和长臂猿科(Hylobatidae,即长臂猿)。人科进一步分化为人亚科(Homininae)和猩猩亚科(Ponginae,即猩猩)。人亚科又进一步分化为人族(Hominini)和大猩猩族(Gorillini,即大猩猩)。人亚科又分为人族(Australopithecina)和黑猩猩族(Panina)。人族最终分化为南方古猿(Australopithecus)和阿法猿(Ardipithecus)。人类大约在250万年前从南方古猿通过能人(Homo habilis)、直立人(Homo erectus)和智人(Homo sapiens)进化而来。
让我们来讨论一下,在过去的250万年里,人类是否可能是通过基因变化从南猿(类人猿)进化而来的。人类的基因图谱已经存在,但尚无南猿的基因图谱。露西是最著名的南猿,其大脑大小与现代黑猩猩相当。因此,让我们假设南猿的基因与黑猩猩的基因相似。人类和黑猩猩的DNA序列因单核苷酸多态性(SNPs)而相差约1.23%,单核苷酸多态性是指DNA序列中单碱基对的变化。如果考虑到基因组中碱基对的插入和缺失(indels),总的差异就会增大。嵌合体是指在一个物种中存在而在另一个物种中不存在的DNA片段。,这可能导致基因组中额外3%的差异。总的来说,虽然人类和黑猩猩的DNA序列有98-99%的相同之处,但剩下的1-2%的差异,再加上基因调控的变化,造成了两个物种在身体、认知和行为上的显著差异。

图3.12.我们是从类人猿进化而来的吗?
众所周知,黑猩猩的突变率约为每代每1亿个碱基对1个突变,与人类的突变率相当。如果我们假定南猿一代人的生活时间为25年,那么250万年中将经历10万代人。在此期间,总突变率为0.1%(10万/1亿)。这一变异率仅占人类与黑猩猩基因差异的10%。因此,南猿似乎不太可能在250万年内进化成人类。这一估计假设所有的突变都是有益的,尽管大多数突变都是有害的。
这一论点也可以通过考虑随机基因突变对密码子的改变来研究。人类和黑猩猩都有大约2万至2.5万个编码蛋白质的基因。由于替代剪接和翻译后修饰,每个基因可以产生多种蛋白质变体,从而产生了大约8万至10万种独特的功能蛋白质。人类蛋白质中的氨基酸数量从20个到33000个不等。假设人类和黑猩猩之间有1%的基因不同,并且两个物种都有20000个蛋白质编码基因,每个蛋白质平均含有100个氨基酸,那么我们预计黑猩猩的每个蛋白质都需要一个氨基酸变异才能与人类的对应蛋白质相匹配。
要想在黑猩猩DNA中发生这些突变,就必须避免在64个可能的密码子中将密码子突变为终止密码子(UAA、UAG、UGA),因为这种变化会导致蛋白质3200失去功能。在不突变为终止密码子和黑猩猩自身密码子的情况下,在20000个蛋白质中实现1%突变率的概率为(60/64)20000=10-561。即使不考虑移帧突变(核苷酸的插入或缺失),这个概率也是非常低的,几乎不可能随机发生。这一论证表明,宏观进化变化,例如从澳洲原始人到人类的转变,几乎不可能通过随机突变发生。
e.智能设计
智能设计通常被认为是创世论的同义词,是一种科学理论,认为宇宙和生物体最好由智能原因而非自然选择或随机过程等非指导过程来解释。与智能设计有关的一个著名案例是2005年在美国宾夕法尼亚州多佛市举行的联邦法院审判。这次审判的起因是家长们提起诉讼,声称在公立学校教授智能设计违反了宪法。家长们认为,智能设计本质上是宗教性的,在公立学校教授智能设计违反了《美国宪法》的“设立条款”,该条款规定政教分离。
在审判过程中,智能设计和进化论的支持者分别提出了自己的论点。代表智能设计的著名人物是生物化学家迈克尔-比赫(MichaelBehe),他断言生物体的复杂结构无法仅用自然选择来解释,并提出了某些特征是由智能原因塑造的可能性。
然而,法院驳回了Behe和其他智能设计支持者的论点,转而接受了进化论支持者的立场。法官裁定,教授智能设计是违宪的,因此在多佛公立学校教授智能设计是非法的。
这一裁决的主要问题在于法院不加批判地接受了进化论支持者的论点和相关科学论文。这些论文含蓄地假定生命是随机产生的,并将基因对环境的适应错误地理解为进化的证据。然而,正如表3.2所概述的,进化论只适用于现存的生物体,无法解释生命的起源。此外,进化理论仅仅描述了已经嵌入遗传密码中的基因的行为。然而,法院在裁决中却没有考虑这些科学事实,这引起了人们对裁决公正性的极大关注。
威廉-佩利(William Paley)是18世纪的哲学家,他是这一论点的奠基人,他用钟表匠的比喻对此进行了著名的说明。佩利认为,正如手表的复杂性意味着,生命和宇宙的复杂性也意味着造物主的神圣。他的观点为现代智能设计理论奠定了基础。智能设计的关键概念包括特定复杂性、不可还原复杂性和微调。第1章和第2章展示了微调的几个例子。现在,让我们详细研究特定复杂性和不可还原复杂性。
i.指定复杂性
特定复杂性是智能设计中的一个关键概念,它认为自然界中的某些模式既高度复杂,又有特定的排列方式来实现特定的功能,这表明是有目的的设计。与随机复杂性不同,特定复杂性不仅错综复杂,而且有序地实现特定的结果。这种双重特性表明,这种模式不可能仅仅是偶然产生的。
DNA的结构就是特定复杂性的例子之一。DNA中的核苷酸序列非常复杂,即使是单链也有数十亿种可能的组合。这种复杂性确保了这种排列不是简单、随机过程的结果。DNA复制和修复机制进一步凸显了其复杂性。这些过程涉及多种蛋白质和酶的协调工作,以准确复制和维护遗传信息。核苷酸序列不仅复杂,而且具有高度特异性,因为它编码合成蛋白质的精确指令。DNA序列中的每个基因都对应着一种特定的蛋白质,即使是序列中的微小变化也会对所产生的蛋白质的功能产生重大影响。DNA还包含控制基因何时何地表达的调节元件,这又为其功能增加了一层特异性。
在DNA中观察到的特定复杂性不太可能是通过随机突变和自然选择等非定向过程产生的。相反,它表明,对于如此复杂和功能特殊的信息的起源,智能原因是一个更合理的解释。
特定复杂性的另一个例子是细菌鞭毛,这是一种鞭状的电动结构,被某些细菌用于运动。下面将详细介绍为什么细菌鞭毛被认为是特定复杂性的一个例子。
细菌鞭毛由大约40种不同的蛋白质组成,这些蛋白质构成了丝状体、钩状体和基质体等不同的组成部分。基体本身的功能就像一个旋转发动机,包括转子、定子、传动轴和螺旋桨。鞭毛要发挥作用,所有这些部件都必须存在并正确组装。缺少其中任何一个部件都会导致鞭毛失去功能,这也凸显了鞭毛的复杂性。
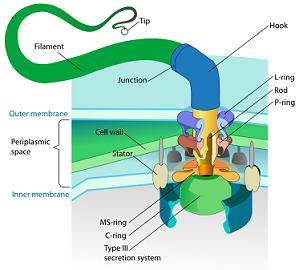
图3.13.细菌鞭毛
鞭毛的各组成部分必须以非常,才能发挥作用。蛋白质必须按照精确的顺序排列,它们的形状也必须完全吻合,就像精心设计的机器部件一样。鞭毛不仅复杂,而且具有高度特定的功能:推动细菌。它的运行速度极快,可以改变方向,而且能效很高,所有这些都表明它的设计是有目的的。
随机突变和自然选择无法充分解释细菌鞭毛的特定复杂性。这种高度集成的功能系统偶然出现的可能性极低。此外,由于鞭毛的中间形态很可能是无功能的,因此传统的逐步改进的进化途径似乎是不可信的。
鞭毛也是不可还原复杂性的典范,它是特定复杂性的一个子集,下文将详细介绍。其论点是,鞭毛的所有部分都是其功能所必需的,因此,它不可能像达尔文进化论所认为的那样,是通过连续的、轻微的修改进化而来的。
ii.不可还原的复杂性
不可降低的复杂性“是生物化学家迈克尔-比赫提出的一个概念,认为某些生物系统过于复杂,不可能是通过循序渐进的改造进化而来的。这些系统,如细菌鞭毛或血液凝固级联,由多个相互依存的部分组成,所有这些部分都必须存在并发挥作用,系统才能运转。任何一个部分的缺失都会导致系统无法运行。这种错综复杂、,相互依存的结构表明存在着智能设计者,因为仅靠自然选择和随机变异是无法解释它们的。这一概念对传统的进化论提出了挑战,并支持自然界中存在有目的的设计这一观点。
视觉循环就是不可简化复杂性的一个例子,它是眼睛中的一个生化过程,将光线转换成电信号,从而实现视觉。该系统由多个相互依存的部分组成,这些部分必须全部存在并发挥作用,该过程才能有效运作。如果缺少任何一个部件或其功能失效,整个视觉循环就会失效,这也说明了不可还原复杂性的概念。视觉周期的主要组成部分包括光感受器(视杆细胞和视锥细胞)、罗得普素、视蛋白、视网膜、信号转导途径和神经处理。
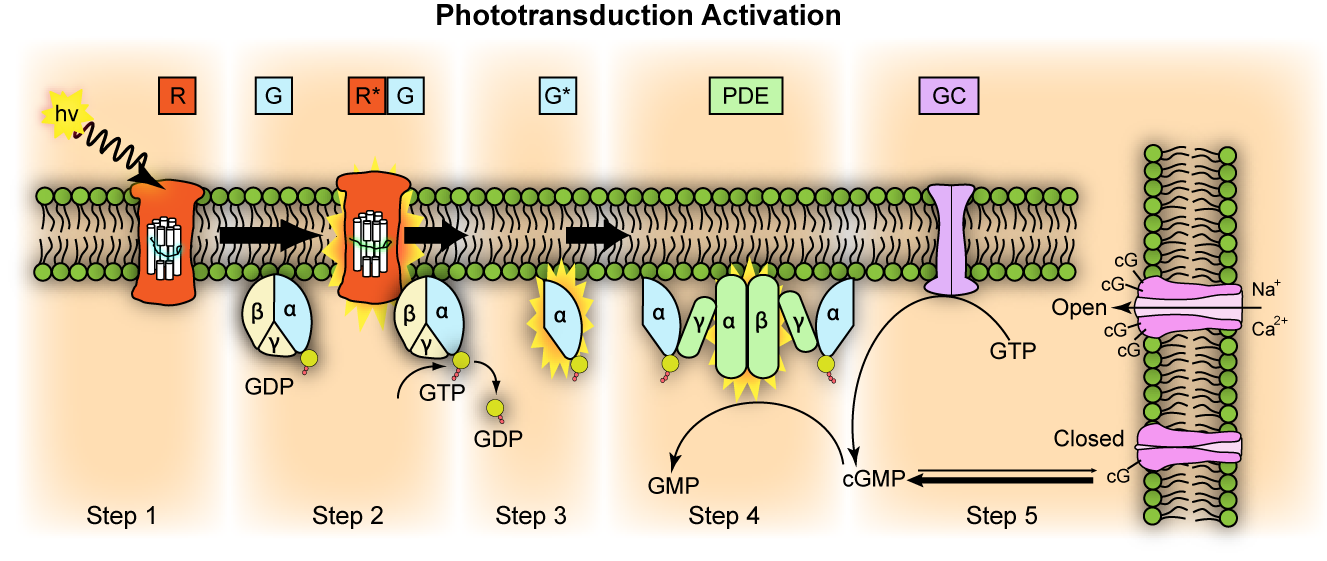
图3.14.视觉周期的分子步骤
感光器是视网膜上检测光线的细胞。视杆细胞负责弱光视觉,而锥体则检测颜色。每个光感受器都含有称为光敏色素的光敏分子,主要是视杆细胞中的视紫红质。视杆细胞中的这种光色素由一种叫做视蛋白的蛋白质和一种叫做视网膜的感光分子组成。锥状体含有不同的视蛋白,能对不同波长的光做出反应,从而产生色觉。视网膜是维生素A的衍生物,在吸收光线时会改变形状。这种形状变化会激活视蛋白,从而启动视觉传导级联。激活的视蛋白又会激活一种叫做转导蛋白的G蛋白。转导蛋白激活磷酸二酯酶(PDE),从而降低细胞中的环磷酸腺苷(cGMP)水平。cGMP的减少会关闭感光细胞膜上的离子通道,导致细胞超极化并产生电信号。电信号通过双极细胞传递到神经节细胞,神经节细胞通过视神经将信号传给大脑。大脑处理这些信号,形成视觉图像。
视觉周期的每个组成部分都是相互依存的。光感受器、视网膜红蛋白、视网膜转导蛋白、PDE和离子通道必须全部存在并正常工作,才能产生视觉。去掉任何一个组件都会导致系统失效。我们可以说,这样一个复杂的系统不可能是通过一系列微小的渐进式变化进化而来的,因为没有所有成分的中间阶段将是无功能的,因此不会受到自然选择的青睐。视觉周期所涉及的错综复杂的生化途径和精确的分子相互作用凸显了视觉所需的复杂性和特异性。其组成部分的相互依存性和所涉及的生化过程的复杂性表明,这一系统不可能通过无定向的进化过程产生,而是指向一个智能设计者,即神圣的造物主。
计算机程序的可视化循环有助于说明其复杂性和相互依存的过程。下面是一个使用python的概念性类比:
用计算机程序编写的视觉循环
#初始化:建立视觉周期的环境,包括感光器(视杆细胞和视锥细胞)
class VisualCycle:
def __init__(self):
self.photoreceptors = {'rods': [], 'cones': []}
self.initialize_photopigments()
self.signal_pathway_active = False
#用户输入:检测进入的光线,启动光敏色素激活程序
def detect_light(self, light_wavelength):
if light_wavelength in visible_spectrum:
self.activate_photopigment(light_wavelength)
#触发事件:改变视网膜的形状并激活视蛋白,然后触发信号转导途径
def activate_photopigment(self, wavelength):
retinal = self.change_retinal_shape(wavelength)
opsin = self.bind_retinal_to_opsin(retinal)
self.start_signal_transduction(opsin)
#事件处理:激活转导蛋白和PDE,导致cGMP水平下降,关闭离子通道,产生电信号
def start_signal_transduction(self, opsin):
self.signal_pathway_active = True
transducin = self.activate_transducin(opsin)
pde = self.activate_pde(transducin)
self.regulate_cGMP_levels(pde)
self.generate_electrical_signal()
#信号处理:根据cGMP水平调整离子通道,以促进电信号的产生
def regulate_cGMP_levels(self, pde):
cGMP_level = self.reduce_cGMP(pde)
self.adjust_ion_channels(cGMP_level)
#信号输出:创建并向大脑传输电信号
def generate_electrical_signal(self):
if self.signal_pathway_active:
electrical_signal = self.create_signal()
self.transmit_signal_to_brain(electrical_signal)
#网络通信:通过双极细胞和神经节细胞处理和转发信号,最终通过视神经发送信号
def transmit_signal_to_brain(self, signal):
bipolar_cells = self.process_signal_with_bipolar_cells(signal)
ganglion_cells = self.forward_signal_to_ganglion(bipolar_cells)
optic_nerve = self.send_signal_via_optic_nerve(ganglion_cells)
self.visual_perception(optic_nerve)
#最终输出:大脑对信号进行解码和处理,生成视觉图像
def visual_perception(self, optic_nerve):
visual_cortex = self.decode_signal(optic_nerve)
self.render_image(visual_cortex)
这个比喻说明了视觉周期中相互依存的步骤和复杂性,就像一个计算机程序,有多个功能和事件处理程序共同实现特定的输出。如果我们错过了其中任何一个步骤,或者使用它们的顺序不对,就无法达到预期的效果。
视觉周期可以用计算机程序来表示,这表明眼睛是智能设计的。眼睛的设计蓝图与位于11号染色体上的PAX6基因有关,该基因在眼睛的发育过程中起着至关重要的作用。
iii.关于智能设计的著名书籍
《Evolution:A Theory in Crisis》(Michael Denton,1985年):Denton对达尔文进化论进行了批判,认为仅靠自然选择无法充分解释生物系统的复杂性。丹顿从分子生物学和古生物学等不同领域提出证据,强调进化论中的漏洞和不一致之处。他认为,在生物体中观察到的错综复杂的结构和功能表明,这是智能设计的结果,而不是随机突变和选择的结果。该书挑战了普遍的科学共识,并提出需要另一种解释来说明生命的起源和多样性。
《Darwin's Black Box:The Biochemical Challenge to Evolution》(Michael J. Behe,2006年):在这本开创性的著作中,迈克尔-比赫提出了“不可还原的复杂性”这一概念,认为某些生物系统(如细菌鞭毛)过于复杂,仅靠自然选择无法进化。贝赫认为,这些系统最好用智能设计来解释。该书对达尔文进化论是否足以解释分子水平上错综复杂的生命机制提出了质疑,并在科学界和哲学界引发了激烈的争论。
《Darwin on Trial》(Phillip Johnson,2010年):本书批判了达尔文进化论的科学基础。身为法学教授的约翰逊以法律分析家的眼光审视了进化论的证据。他认为,自然选择和随机变异并不能充分解释生命的复杂性。约翰逊认为,对达尔文主义的支持大多基于哲学自然主义,而非实证科学。他对科学界不愿考虑其他解释(如智能设计)提出质疑,并呼吁对生命起源进行更开放的讨论。该书在推广智能设计和质疑达尔文理论在生物学中的主导地位方面很有影响力。
《Signature in the Cell:DNA and the Evidence for Intelligent Design》(Stephen C. Meyer,2010年):本书探讨了生命的起源和DNA中编码的信息。迈耶认为,DNA中复杂而具体的信息最好由智能原因来解释,因为自然主义过程无法解释此类信息的起源。他以错综复杂的遗传信息为基础,详细论证了智能设计,指出生命的起源是有目的的创造,而不是随机过程。
《Darwin Devolves:The New Science About DNA That Challenges Evolution》(Michael J. Behe,2020年):贝赫的另一本著作认为,最近的基因发现破坏了传统的达尔文进化论。他断言,虽然自然选择和随机突变可以解释微小的适应性,但它们无法解释细胞内分子机制的复杂性。他提出了“蜕变”的概念,即突变导致遗传信息的丢失,而不是创造出新的有益性状。比赫认为,这些遗传局限性表明了智能设计者的必要性,挑战了传统的进化论框架,并提出智能设计为生命的复杂性提供了更合理的解释。
《The Mystery of Life's Origin:Reassessing Current Theories》(Charles B. Thaxton 等,2020年):这部开创性的著作批判了关于生命起源的各种自然主义理论,并提出了智能设计这一更为合理的解释。他们认为,前生物化学和生命从非生命中的形成,用智能原因来解释更合理。该书讨论了当代生命起源理论的缺点,并介绍了智能设计作为一种科学上可行的替代方案,为现代智能设计运动奠定了基础。
《The Design Inference:Eliminating Chance through Small Probabilities》(William A. Dembski 与 Winston Ewert,2023年):本书为探测自然界中的设计奠定了理论基础。他们探讨了探测智能设计的数学框架。作者提出了一个论点,即表现出特定复杂性的复杂系统最好由智能原因而非随机过程来解释。他们介绍了“特定复杂性”的概念,即复杂性与独立给出的模式相结合。书中运用概率论说明,自然界中的某些模式不可能是偶然产生的。通过严谨的分析,登布斯基和埃沃特认为,承认设计是一种合法的科学实践,并为区分生物系统中的设计和偶然。
f.粒子物理学与创造
在上一节中,我们通过讨论生命的基本组成元素,包括氨基酸、核糖核酸、蛋白质、脱氧核糖核酸和细胞,探索了生命的起源。这些组成部分都是由原子构成的,我们默认原子是自然存在的。原子由基本粒子组成。在本节中,我们将仔细研究这些粒子的起源,探讨它们是自发产生的,还是通过有目的的过程形成的。
根据粒子物理学标准模型,宇宙中的所有物质都由17种基本粒子组成。其中包括6个夸克、6个轻子、4个规玻色子(胶子、光子、Z玻色子和W玻色子)以及希格斯玻色子。每种粒子都有特定的属性,如质量、电荷和自旋,并且每种粒子都在粒子相互作用中发挥着独特的作用,就像细胞中的细胞器发挥着不同的功能一样。
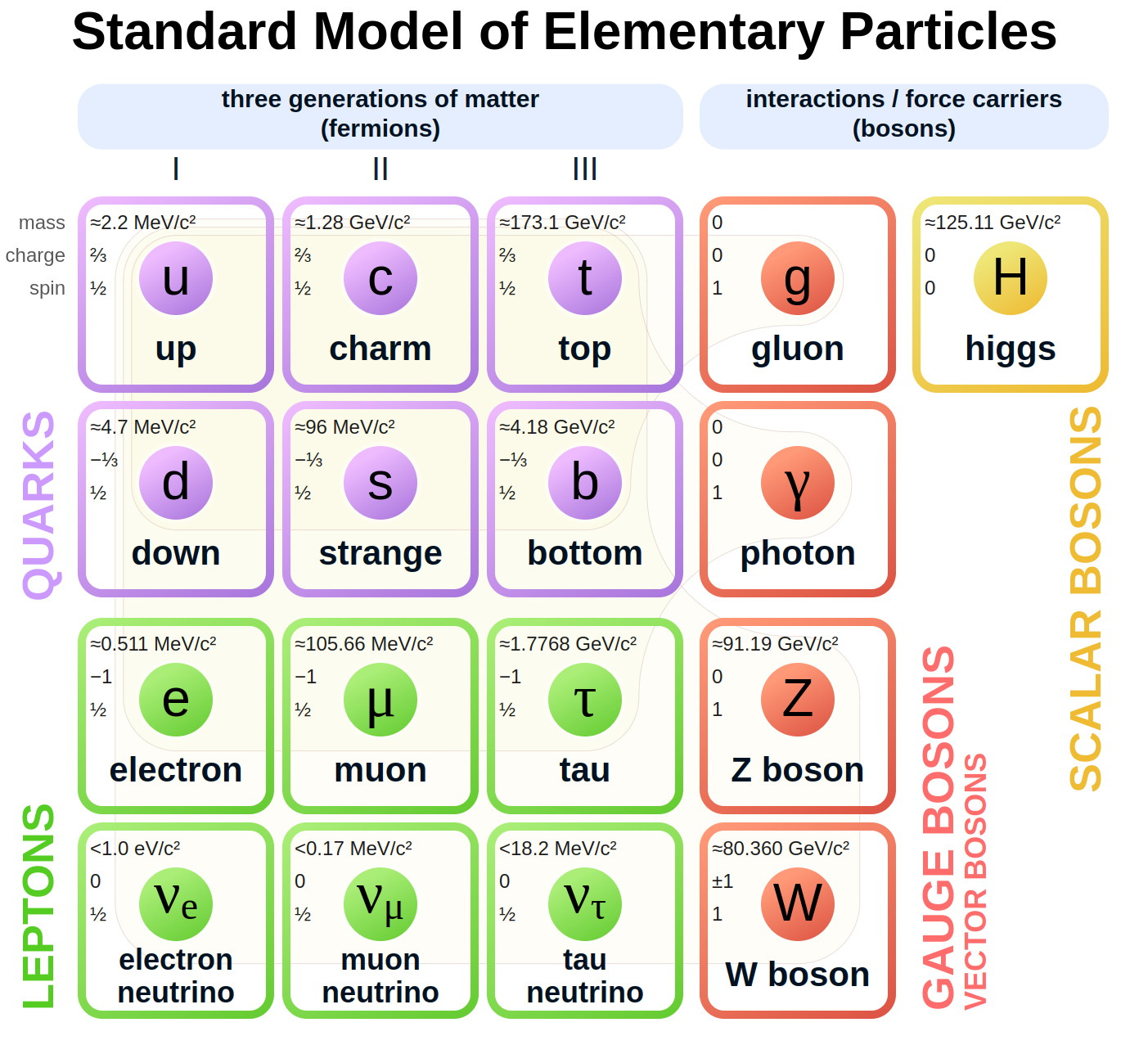
图3.15.标准模型的基本粒子
夸克是物质的基本组成部分,是形成质子和中子的关键。质子由两个上夸克和一个下夸克组成,而中子则由一个上夸克和两个下夸克组成。夸克通过强力结合在一起,并以胶子为媒介。引力或电磁力会随着距离的增加而减弱,而夸克之间的强力则不同,它随着夸克的移动而增加,随着夸克的靠近而减弱,从而保持一定的距离。夸克可以在粒子相互作用过程中改变类型,例如在β衰变过程中,中子通过将下夸克转换为上夸克而变成质子。
量子玻色子是介导自然界基本力的基本粒子。它们包括电磁力的光子、弱力的W和Z玻色子以及强力的胶子。每个规玻色子都与特定的场相关联,并携带粒子间的力。它们对于解释量子层面的相互作用、管理粒子如何相互作用并结合在一起形成物质至关重要。
希格斯机制是一个解释基本粒子如何获得质量的过程。它涉及希格斯场,一种贯穿宇宙的能量场。当粒子与希格斯场相互作用时,它们就会获得质量,就像物体在介质中运动时会遇到阻力一样。希格斯玻色子是一种与希格斯场相关的粒子,于2012年被发现,证实了这一理论。如果没有希格斯机制,粒子将仍然是无质量的,宇宙将缺乏形成原子、生物体、行星和恒星所必需的结构
粒子物理学的运行达到了令人难以置信的先进和复杂水平,为我们提供了有关宇宙本质和起源的深刻见解。这促使我们提出以下基本问题以及其他许多问题:
- 具有如此精确特性的17种基本粒子是如何产生的?
- 规玻色子是如何获得力中介特性的?
- 希格斯机制是如何产生的?
- 贝塔衰变机制是如何产生的?
- 如何用数学方法描述基本粒子的特性?
如果上述问题的答案纯粹是随机过程的结果,那么我们所知的世界可能就不存在了。例如,如果缺少哪怕一个基本粒子,如果希格斯机制尚未建立,或者如果基本粒子的质量和自旋值稍有不同,中子、质子和电子就无法结合在一起。这将导致所有物质的坍塌,使任何事物--包括人类--的形成成为不可能。宇宙基本结构的这种微调精度体现了粒子物理学领域的“不可还原复杂性”概念,而这一原理通常与智能设计有关。
基本粒子形成物质的过程可以比作多细胞生物体中细胞和细胞器的形成过程。正如特定的细胞和细胞器各自具有不同的作用和特性,有助于生物体的复杂功能一样,基本粒子也具有精确的特性,能够形成原子、分子,并最终形成所有物质。这种平行关系强调了自然世界固有的复杂性和意向性--无论是在活细胞的微观层面,基本粒子的亚原子领域,还是在生物体、恒星和星系的宏观层面。
基本粒子的形成及其相互作用可以用量子力学的数学方程精确描述,这一事实表明,它们是有意的数学设计的结果,而不仅仅是偶然的。否则,我们就不得不假定基本粒子具有智慧,能够自行决定形成物质和与其他粒子相互作用所需的质量、电荷和自旋的精确值。然而,我们知道事实并非如此,因为基本粒子没有意识,也没有对量子力学的内在理解。
在生物系统和粒子物理学中观察到的错综复杂的设计和协调有力地表明,存在着潜在的智能和有目的的创造--这是智能设计的标志--而不是一系列随机发生的现象。
g.外星人与创世
几十年来,外星人或外星生命的可能性一直令科学家和公众着迷。宇宙浩瀚无边,每个星系都有数十亿颗恒星,甚至可能有更多的行星,因此,如果生命自发产生,那么在其他地方也可能存在生命,这在统计学上似乎是可信的。一个星系中的地外文明数量可以通过德雷克方程估算出来:N=R*×fp×ne×fl×fi×fc×L其中,N是高级文明的数量,R*是恒星形成率,fp是拥有行星的比例,ne是支持生命的行星数量、fl是有生命发展的行星的比例,fi是有智慧生命进化的行星的比例,fc是可以发送信号的文明的比例,L是文明可以通信的时间长度。如果每个参数都取一个合适的值,一个星系中的文明数量估计约为2。

图3.16.外星人存在吗?
搜寻地外智能项目(SETI)始于1960年。这些项目利用各种方法和技术扫描宇宙,寻找外星文明的证据。以下是一些重要的SETI项目。
奥兹玛计划是第一个现代SETI实验。它利用射电望远镜扫描头鲸鱼座和伊普西隆二号星,寻找潜在的地外信号。SETI@home是一个分布式计算项目,利用家用电脑的闲置处理能力。志愿者在个人电脑上安装软件,分析无线电信号,寻找地外智慧生物的迹象。艾伦望远镜阵列是一个专用的射电望远镜网络,旨在持续系统地搜索地外信号。它由多个小型天线组成,共同勘测天空中的大片区域。突破聆听”是迄今为止最全面的SETI项目,旨在勘测100万颗最接近的恒星和100个附近的星系,寻找潜在的信号。快速射电暴项目调查从太空中探测到的神秘快速射电暴,这些射电暴可以让人们了解未知的宇宙现象。激光SETI项目侧重于探测来自地外文明的光学信号,探索通过激光传输进行星际通信的可能性。
尽管利用先进的射电和光学望远镜进行了持续的搜索,但SETI项目未能找到外星智慧生命的确切证据。

图3.17.用于SETI的射电望远镜
如果存在众多的外星文明,它们可能曾经访问过我们,也可能正在访问我们。在这种情况下,他们会使用什么样的太空旅行方式呢?由于宇宙幅员辽阔,使用飞行物(火箭或飞碟)进行太空旅行面临着难以克服的挑战。即使是最近的恒星--半人马座比邻星(Proxima Centauri),也在4.24光年之外,以目前的技术需要数万年才能到达。由于距离遥远,即使是我们的银河系,也不可能在人类的有生之年探索到,更不用说宇宙了。
可能的先进推进方法包括曲速驱动器或虫洞旅行。曲速驱动器是一种超光速太空旅行的理论概念,其灵感来自爱因斯坦的广义相对论。翘曲驱动器由物理学家米格尔-阿尔库比耶尔于1994年提出,包括制造一个“翘曲气泡”,使航天器前面的空间收缩,后面的空间膨胀。这将使航天器相对于外部观察者的移动速度超过光速,而不违反物理定律。关键的挑战在于,这需要具有负能量密度的奇异物质,而这种物质尚未被发现或创造出来。虽然理论上很有希望,但要使曲速驱动器在太空探索中实际使用可行,还需要重大的科学和技术进步。
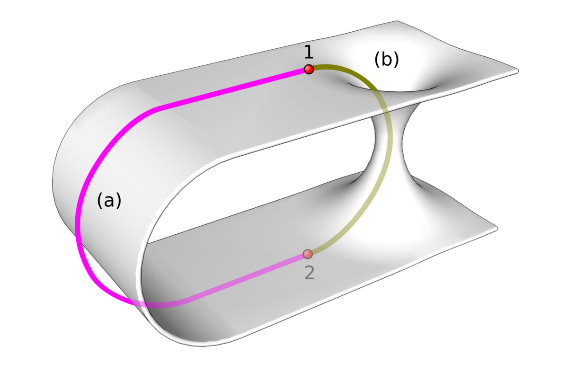
图3.18.虫洞
通过虫洞进行太空旅行是一个理论概念,涉及连接宇宙中遥远点的时空捷径。根据爱因斯坦广义相对论的预测,虫洞或爱因斯坦-罗森桥有可能实现瞬间穿越遥远的宇宙距离。为了实际使用,可穿越的虫洞需要稳定,理论上需要具有负能量密度的奇异物质,以防止坍塌。尽管虫洞是一个流行的科幻小说故事,但它仍然是一种推测,没有实验证据。如果可行,它们将彻底改变太空旅行,使人们能够探索遥远的星系,并将旅行时间从数年缩短到片刻。然而,要使这一概念成为现实,还需要在科学和技术上取得重大突破。
通过超空间或大体积进行瞬移可能是绕过传统三维空间实现瞬间穿越遥远距离的另一种方法。超空间指的是在我们熟悉的三维空间和一维时间之外的额外维度或一系列维度,为穿越宇宙结构提供了一条捷径。同样,“体”也是弦理论中的“鹤膜宇宙学”等理论中使用的术语,在这些理论中,我们的宇宙被设想为被称为“体”的高维空间中的“鹤膜”。在这些理论中,远距传物是指通过这些高维空间的移动,瞬间重新出现在我们宇宙中的另一个位置。兰德尔-孙德鲁姆模型(Randall-Sundrum model)等理论框架提出,存在这样的高维空间,可以实现时空捷径。如果这种维度存在并且可以进入,那么就有可能利用它们进行远距传物,从而避免相对论旅行的限制,并有可能使超光速旅行变得可行。

图3.19.瞬移
如果生命像德雷克方程假设的那样自发产生,那么宇宙中的地外文明总数将达到约4000亿个(2000亿个星系中每个星系有2个文明)。地球上的生命大约始于40亿年前。现在,设想有1%的地外文明比我们早100万年开始,并遵循类似的进化路径。在这种情况下,它们的文明将比我们的文明先进100万年。有了如此显著的领先优势,他们可能已经开发出了先进的远程传送技术,使他们能够像我们访问邻居一样轻松地前往宇宙中的任何地方。如果一个这样的文明的人口是10亿,那么外星人的总数将是五亿(1018)。如果其中只有1%的外星人能够每10年访问地球一天,那么地球上每天就会挤满大约10万亿个外星人--是目前人类人口的1000倍。然而,我们没有观察到任何外星人存在的证据。我们如何解释这种明显的矛盾呢?
这个问题被称为“费米悖论”(Fermi Paradox),以恩里科-费米(Enrico Fermi)的名字命名。答案可能是:(i)德雷克方程中的假设(进化)是错误的,或者(ii)先进文明可能使用我们目前的方法无法检测到的技术,或者故意避免被检测到。如果外星人既不是细菌也不是隐形生物,那么他们的存在很可能早就以某种方式向我们揭示了。然而,我们尚未探测到他们存在的任何证据,这一事实表明德雷克方程中的进化假设很可能是不正确的。
h.生物和造物的本能
计算机由三个主要部分组成:硬件、软件和固件。固件是编程到ROM或UEFI中的专用软件,为特定硬件提供关键控制,是硬件和软件之间的中介。它对于系统启动、管理硬件操作和确保设备功能至关重要。
计算机中的固件和生物体中的本能有一个重要的相似之处:两者都是内在的、预先编程的系统,管理着基本功能。固件对操作进行初始化和管理,确保开机后的正常功能。同样,本能也是一种自然的、与生俱来的行为模式,指导着进食、交配和逃离危险等生存活动。这两种系统都是在无意识输入的情况下自动运行,为有效运作和环境响应提供基础指导。从本质上讲,固件之于计算机,就如同本能之于生物体--一个嵌入式的、预先配置好的系统,对基本操作和生存至关重要。正如固件是由计算机设计师嵌入ROM一样,本能也是由神圣的造物主嵌入生物体的大脑和神经系统。让我举几个本能的例子来说明这个概念。
i.泥瓦匠蜂的筑巢
在让-亨利-法布尔的著作《泥瓦匠蜂》(《昆虫记》的一部分)中,他描述了泥瓦匠蜂复杂的筑巢过程。这些蜜蜂选择一个合适的平面(通常是石头)开始筑巢。它们收集泥浆和小石子,一丝不苟地为后代建造巢室。雌蜂将泥团运到现场,将它们塑形并压实,形成安全的细胞壁。然后,雌蜂采集花蜜和花粉,为每个细胞提供营养,产下一粒卵,再用更多的泥土将其密封。这个过程不断重复,最终形成了一系列排列整齐、用卵石加固的泥室,保护着正在发育的幼虫。法布尔的观察凸显了这些独居蜜蜂非凡的精确性和勤劳。
他描述了一个实验,他将一个未完成的巢与一个已完成的巢进行了交换。石匠蜂在发现自己未完工的巢被完工的巢取代后,表现出了一种有趣的行为。蜜蜂没有继续在新巢上工作,而是继续建造,就像没有发生任何变化一样。它不承认已完成的巢是自己的作品,而是坚持自己的习惯动作,带着泥巴继续建造。
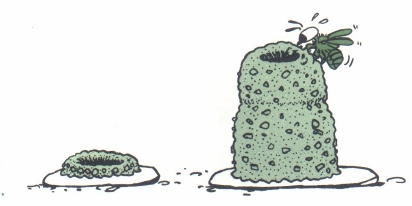
图3.20.泥瓦匠蜂在已建成的上筑巢
这个实验说明了蜜蜂行为的本能性和程序性,其驱动力是内部的一连串动作,而不是巢穴状态的视觉线索。
法布尔做了一个相反的实验,将一个已完成的泥蜂巢与一个未完成的泥蜂巢对调。他观察到,当石匠蜂回到现场,发现已完成的巢被未完成的巢取代时,它并没有继续在新的未完成的巢上工作。相反,蜜蜂似乎很困惑,花了一些时间检查被改动过的巢,但最终没有继续建造。然后,它又开始了下一个动作,即使蜂蜜已经溢出,也要把它填满。这种行为表明石匠蜂对其特定巢穴的强烈依恋,以及适应环境中意外变化的困难。这个实验还突出了石匠蜂筑巢过程的本能性质。

图3.21.泥瓦匠蜂向未完工的巢中灌蜜
法布尔还做了另一个有趣的实验。泥瓦匠蜂先把花蜜装满巢穴,然后转180度,掸掉腿上和身上的花粉。如果它在掸花粉时受到干扰,就会飞走,等待威胁过去。回到巢中后,它又从头开始。即使它的花蜜窝里什么也没有,也要给它装满花蜜。这个实验表明,蜜蜂本能地遵循固有的采蜜程序,它们的行动顺序是无法改变的。
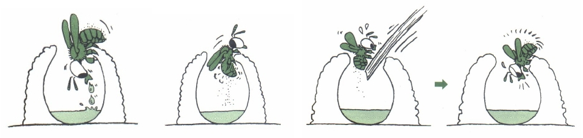
图3.22.石匠蜂受到干扰时的行为
泥瓦匠蜂完成筑巢后,会在巢中装满花蜜和花粉,在上面产卵,然后封住巢顶。法布尔进行了另一项实验:在一个巢的顶部贴上纸,在另一个巢的顶部放上一个纸锥。他观察了孵出的泥瓦匠蜂的行为。在粘贴了纸的巢穴中,蜜蜂用它强壮的下颚顺利地切开了顶部。对于贴有纸筒的巢穴,蜜蜂切开了顶部,但不知道下一步该做什么。它本想看到开阔的天空,却被纸筒迷惑了,没有尝试刺穿纸筒,最终死亡。

图3.23.用纸粘贴并用纸筒覆盖的蜂巢
上述所有实验都表明,泥瓦匠蜂的行为具有本能和程序化的性质,是由其遗传密码中的内部行动序列驱动的。
ii.筑巢
编织鸟以其复杂精致的巢穴而闻名,它巧妙地将草叶和其他植物材料编织成复杂的结构,展示了非凡的工艺和本能的工程学。

图3.24.纺织鸟的巢
南非博物学家和诗人尤金-马莱斯(Eugène Marais)对纺织鸟进行了引人入胜的实验,研究它们的筑巢行为和本能的作用。马莱斯的目的是了解织女鸟复杂的筑巢技能是纯粹的本能,还是涉及学习行为。
马莱斯在与自然环境隔离的环境中饲养织金鸟,确保它们不会接触到其他鸟类或筑巢活动。他观察了这些被隔离的鸟从孵化到成熟的整个过程,确保它们四代都没有机会向其他织女鸟学习。对于第五代,马莱斯提供了与野生织金鸟相同的筑巢材料,如草和树枝。尽管从未见过鸟巢或其他鸟类筑巢,但这些与世隔绝的织女鸟开始筑巢,筑出的巢与野生织女鸟几乎一模一样。它们展示了同样复杂的编织技术、打结方法和整体结构。这些与世隔绝的鸟类筑的巢显示出与同类一致的典型设计特征,表明它们的筑巢技能是与生俱来的,而不是通过观察或模仿学会的。
马莱斯得出结论,纺织鸟复杂的筑巢行为是由本能驱动的。这种与生俱来的行为编码在它们的大脑和神经系统中,使它们能够在没有经验或学习的情况下建造复杂的巢穴。这些与生俱来的行为是有目的的设计,并通过DNA世代相传。
iii.鹦鹉螺的形成贝壳
鹦鹉螺是一种以美丽独特的贝壳闻名的海洋软体动物。其贝壳形状遵循精确的对数螺旋线。鹦鹉螺贝壳的形成是本能的又一惊人例证,涉及生物和化学过程的复杂相互作用,这些过程精密协调以产生其独特结构。
这一过程始于鹦鹉螺还是卵内胚胎时。在此阶段,最初的壳体(称为原壳)开始形成。这个初始腔室体积较小,为后续壳体生长奠定基础。包裹壳体的特殊组织——外套膜,会分泌以方解石(CaCO3)形式的碳酸钙层。外套膜细胞从海水中提取钙离子,并与碳酸根离子结合形成碳酸钙。外套膜还分泌一种由蛋白质和多糖组成的有机基质,作为碳酸钙沉积的支架。这种基质有助于控制方解石晶体的形状和取向,确保贝壳的强度和耐用性。

图3.25.显示对数螺旋图案的鹦鹉螺壳
随着鹦鹉螺的生长,它会定期在其壳上添加新的腔室。每个新腔室都比前一个更大,以容纳不断增大的鹦鹉螺。鹦鹉螺在壳内向前移动,并用称为隔板的墙壁密封较旧的腔室,从而形成一系列逐渐增大且相互连通的腔室。一个名为“ siphoncle ”的特殊器官贯穿贝壳的所有腔室。这种管状结构调节腔室内气体和液体的含量。通过控制气体(主要是氮气)和液体的水平, siphoncle 帮助鹦鹉螺调节浮力,使其能在水柱中上下移动。外壳的最外层称为外壳膜,是一层有机层,保护下方的碳酸钙层免受溶解和物理损伤。外壳膜下方是方解石层,呈珍珠层或棱柱状结构,为外壳的虹彩和强度做出贡献。
碳酸钙的分泌、腔室的形成以及通过 siphon 调节浮力的复杂协调,表明这是一个“全有或全无”的系统,其复杂性超出了渐进进化的能力。化石记录中缺乏明确的过渡化石,加上鹦鹉螺被称为“活化石”,暗示其突然出现,并表明其复杂的壳体形成指向有目的的创造而非无方向的进化。鹦鹉螺不具备数学或生物化学知识;因此,其对数螺旋壳形的精准形成、复杂的生物化学分泌调控,以及浮力系统的无缝整合,并非随机过程的结果。相反,这些特征表明存在一个预先编程的遗传蓝图,使鹦鹉螺能够以惊人的精度构建其复杂壳体,进一步强化了有目的设计而非无指导进化的观点。
i.自然和创造中的数学
“数学是上帝书写宇宙的语言”。-伽利略-伽利莱
数学模式和原理在自然界中比比皆是,包括黄金分割率、黄金分割角、斐波纳契数列、对数螺旋和分形。
- 黄金分割率通常用希腊字母φ表示(=(a+b)/a=a/b),是一个无理数,约等于1.618。当两个量的比值与它们的和与两个量中较大的的比值相同时,就出现了黄金比。
- 黄金角是将圆按黄金分割率分成两个弧长的两个半径所夹的角。它是按照黄金分割率分割圆周时产生的两个角(约137.5度)中较小的一个角
- 斐波那契数列是从0或1开始的一系列数字,其中每个数字都是前两个数字之和(例如0、1、1、2、3、5、8......)。
- 对数螺旋是自然界中经常出现的一种自相似螺旋曲线。它的特点是任意一点的切线和径线之间的夹角是恒定的。
- 分形是在不同尺度上具有自相似性的复杂图案。它们通常是通过在一个持续的反馈循环中不断重复一个简单的过程而产生的。
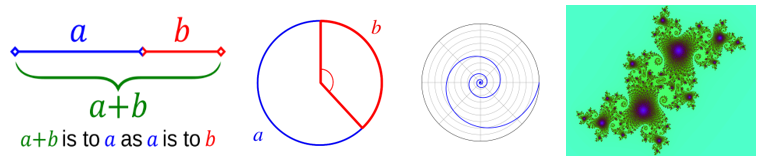
图3.26.黄金分割率、黄金分割角、对数螺旋和分形
让我们来探索一下这些数学原理在自然界中的应用。
植物叶轴是指叶、花或其他植物结构在植物茎上的排列。它是植物学中的一个重要概念,反映了植物最大限度地接触阳光和其他环境资源的方式。叶片的排列遵循斐波那契数列,其中连续螺旋状排列的叶片数是斐波那契数字。可能的叶轴模式有1/2,1/3,2/5,3/8,5/13,8/21等,其中分子和分母构成斐波那契数列。
3/8植轴指的是一种叶片排列模式,每片叶片与下一片叶片之间的距离是围绕茎旋转360度的八分之三。这意味着每一片叶子都与前一片叶子成3/8×360=135度角(称为发散角)。在叶片数量较多的植物中,发散角趋近于黄金分割角137.5度。这种微小的发散角有助于叶片的分布,最大限度地增加阳光照射,减少重叠和遮荫,确保每片叶子都能获得充足的光照和空气。适当的间距可使水分和养分在整个植株中得到最佳分配。
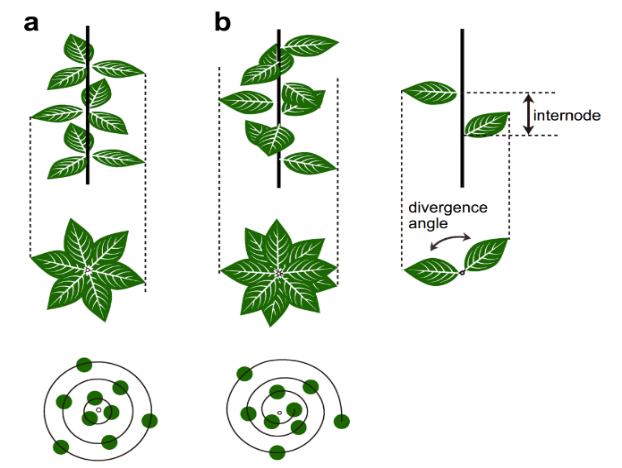
图3.27.2/5植轴(a)和3/8植轴(b)
在许多花卉中也能发现类似的模式。例如,喷嚏草的叶子、枝条和花瓣的数量构成了连续的斐波纳契数。叶片为1、1、2、3、5、8,枝条为1、2、3、5、8、13,花瓣为5、8或8、13。
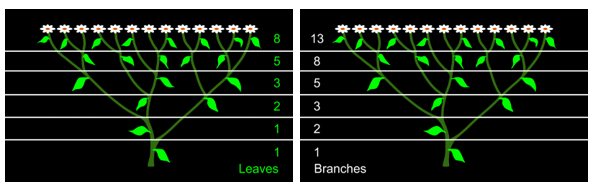
图3.28.喷嚏草的叶子和枝条
不仅是叶子,植物的嫩芽、果实和种子都受斐波那契数列和黄金分割角的支配。
挪威云杉的发芽模式遵循斐波那契数列和黄金分割角的原理。每一个新芽的萌发角度都与前一个新芽的萌发角度约为137.5度(黄金分割角)。
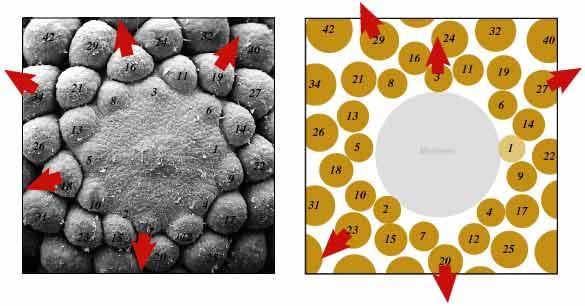
图3.29.挪威云杉的萌芽模式
因此,树枝围绕树干形成螺旋状,其分布与斐波那契数字一致。这种自然模式增强了树木有效收集阳光、水分和养分的能力,支持其生长和健康。雏菊的花朵排列也体现了斐波纳契模式和黄金分割角。花瓣和种子按照斐波那契数列呈螺旋状排列,每个方向的螺旋数通常对应连续的斐波那契数字,如21和34。此外,连续花瓣或种子之间的发散角近似黄金分割角。如果螺旋以黄金分割角缠绕,就会形成对数螺旋。如果雏菊的小花形成对数螺旋,它们在生长过程中就会保持其形状。对数螺旋具有自相似性,这意味着螺旋的形状即使在扩展过程中也会保持一致。对数螺旋的固有特性使雏菊在整个生长过程中都能保持其整体几何结构。
松果、花椰菜和罗马椰菜中也有类似的图案。松果的鳞片呈复杂的螺旋状排列,遵循斐波那契数字,通常一个方向有8个螺旋,反方向有13个螺旋,每个鳞片的位置都经过精心设计,大致呈黄金分割角。同样,花椰菜的小花在一个方向上缠绕成5个螺旋形,在另一个方向上缠绕成8个螺旋形,反映了相同的数字顺序。在罗马菜花中,小花在一个方向上呈13个螺旋状排列,在另一个方向上呈21个螺旋状排列。
菠萝中的斐波纳契数可以在其眼睛的排列中找到。这些眼睛按照斐波那契数字排列成螺旋状,通常形成三组不同的螺旋。通常情况下,在一个方向上有8个螺旋上升,在相反方向上有13个螺旋上升,有时在另一个方向上有21个螺旋上升,每组螺旋都与连续的斐波纳契数一致。这种模式确保了包装的效率,并最大限度地提高了水果结构的完整性。这种排列使菠萝生长均匀,养分分布均匀,展示了斐波纳契数列在植物生长和发育中的自然应用。

图3.30.植物中发现的斐波那契数列和对数螺旋线
对数螺旋式的生长曲线不仅出现在植物中,也出现在人类和其他动物中。例如,人类的耳廓、耳朵的耳蜗、人类的手指、海马的尾巴、山羊角以及包括鹦鹉螺在内的各种蜗牛的壳。如果这些生长模式不遵循对数螺旋,它们在继续生长的过程中就无法保持其特有的形状,最终会失去其独特的功能和独特的结构完整性。
例如,如果耳蜗的生长模式不遵循对数螺旋,就会严重影响其有效处理声音的能力。对数螺旋可使频率沿其长度方向梯度检测,高频在底部,低频在顶点。偏离这种模式会导致频率检测区域的间距不均匀,从而导致听力受损或难以区分不同的声音频率。这种精确的排列对于耳蜗将声波转化为神经信号、实现准确的听觉感知至关重要。

图3.31.耳蜗、耳朵、海马和手节骨
在自然界中可以发现许多分形图案,包括蕨类植物和树木的分枝图案、蕨类植物叶子的结构、花椰菜、西兰花和罗马椰菜中小花的排列、许多植物的根系以及松果。分形图案也存在于生物系统中。
大到动脉,小到毛细血管,血管的分支都遵循分形模式。分形结构最大限度地扩大了营养物质和气体交换的表面积,同时将泵送血液到全身所需的能量降至最低。分形分支确保每个细胞都能获得充足的氧气和营养物质。此外,血管的分形特性还有助于增强其坚固性和适应性。重复模式可以轻松适应生长和修复,在发生变化或损坏时仍能保持高效循环。
人类的呼吸系统也有分形模式。肺的结构由气管分支成支气管,支气管再分支成更小的支气管,最后形成进行气体交换的肺泡。每个分支都保持着分形模式。这种分形结构最大限度地扩大了进行气体交换的表面积(相当于网球场大小),同时最大限度地缩小了肺部所占的体积。通过遵循分形模式,肺部可以有效地向血液输送氧气并排出二氧化碳,从而优化呼吸功能。

图3.32.在蕨类植物和罗马式中发现的分形
自然界和生物系统中存在的黄金分割角、斐波纳契数列和分形等数学模式,对随机突变和自然选择的观点提出了挑战。例如,金角的最佳叶间距和斐波那契数列在种子排列方面的高效率,都表明这是一种有目的的设计,以最大限度地利用资源。分形在血管和植物根系等结构中的自相似复杂性表明,随机过程无法达到复杂的组织水平。这些结构的复杂性、精确性和普遍性表明,这是一种预先确定的智能设计,而不是无定向的进化过程。
邀请福音
“我观看你指头所造的天,并你所陈设的月亮星宿,
便说:人算什么,你竟顾念他?世人算什么,你竟眷顾他?
你叫他比天使微小一点,并赐他荣耀尊贵为冠冕。
你派他管理你手所造的,使万物,就是一切的牛羊、田野的兽、
空中的鸟、海里的鱼,凡经行海道的,都服在他的脚下。
耶和华—我们的主啊,你的名在全地何其美!”(诗篇8:3-9)
上述《圣经》经文美妙地反映了对造物的敬畏和惊奇,承认诸天的威严和宇宙的复杂设计是造物主的证据。在这些经文中,诗人惊叹于月亮、星星和广阔无垠的天空,这些都是上帝创造的,他认识到创造是有意识、有目的的行为。创造论利用了这种惊奇感,断言大自然中的复杂性和秩序不是随机偶然的产物,而是神圣的造物主有意设计的结果。诗人反思人类的渺小与宇宙的宏伟相比,强调了这样一种信念,即尽管宇宙浩瀚无边,但上帝选择以荣耀和尊贵为我们加冕,让我们掌管他手所造之物。上帝与人类之间的这种深厚关系指向他对我们的深爱,以及他希望我们与他生活在一起的愿望。
在本章中,我想介绍福音,它揭示了上帝的爱和与我们相交的愿望是如何通过耶稣基督实现的,为我们提供了与祂和好并活在祂丰盛恩典中的机会。对于那些仍然难以相信上帝存在的人,我还想介绍帕斯卡尔的赌注。
布莱斯-帕斯卡是17世纪法国哲学家、数学家、物理学家和作家,因其对人性和信仰的哲学思考而闻名于世,尤其是在他的作品《笔谈》中。他提出了一个关于上帝存在的哲学论证,称为“帕斯卡赌注”。帕斯卡尔认为,活得像上帝存在一样是一个理性的决定,因为如果上帝真的存在,信徒就会获得永恒的幸福,而如果上帝不存在,损失则微乎其微。相反,如果一个人活得好像上帝不存在,并且是错误的,那么潜在的损失是巨大的,包括永恒的痛苦,而如果是正确的,获得的则微乎其微。因此,帕斯卡尔得出结论,相信上帝是更安全、更有益的“赌注”。
上帝存在 | 上帝并不存在 | |
相信上帝 | 永恒的欢乐(天堂) | 什么也没发生 |
不上帝 | 永恒的痛苦(地狱) | 什么也没发生 |
表4.1.帕斯卡赌注
到目前为止,我们已经就创造和进化进行了广泛的讨论,承认了上帝的存在。如果你承认这一真理,那么帕斯卡尔的赌注就提出了两个明确的选择:永恒的快乐(天堂)或永恒的痛苦(地狱)。每个人都希望选择第一个选项,没有人愿意选择第二个选项。在这个阶段,你可能会怀疑天堂的存在,但天堂确实存在。在《哥林多后书》中,使徒保罗分享了一段深刻而神秘的经历,从中可以窥见天堂的存在。他写道:
“我认得一个在基督里的人,他前十四年被提到第三层天上去;或在身内,我不知道;或在身外,我也不知道;只有神知道。
我认得这人;或在身内,或在身外,我都不知道;只有神知道。
他被提到乐园里,听见隐秘的言语,是人不可说的。”(哥林多后书12:2-4)
保罗的叙述表明,天堂或“第三层天”是一个无法形容的美丽境界,是神的存在,与我们的尘世经历截然不同。这“第三层天”被认为是天堂的最高部分,是终极灵性现实和与上帝交流的地方。保罗在那里听到的“无法言喻的事”表明,天堂的经历和真理超出了人类的理解和语言。
这段经文让信徒们确信天堂的真实性及其深远、超越的本质,为我们带来了希望,并应许了在我们尘世之外等待着我们的神圣奥秘。保罗的异象有力地证明了天堂的存在,这是上帝为爱祂的人预备的地方。
天堂向任何相信耶稣基督的人敞开大门。耶稣基督来到人间,将人类从罪恶中拯救出来。耶稣是一个历史人物。我们的历史按公元前(B.C.:Before Christ)和公元后(A.D.:Anno Domini,拉丁语,意为“耶和华年”)划分。正如四卷福音书中记载的那样,耶稣在传道期间行了无数神迹,展示了他的神力和慈悲。他医治病人,如治好麻风病人(马太福音8:1-4)和使盲人复明(约翰福音9:1-7)。他还行自然神迹,包括平息风暴(马可福音4:35-41)和在水面上行走(马太福音14:22-33)。此外,耶稣还使死人复活,尤其是拉撒路(约翰福音11:1-44),并使饼和鱼增多,喂饱了成千上万的人(《马太福音》14:13-21)。这些神迹肯定了耶稣作为神子的身份,并给许多人带来了希望和信心。
如果你想信耶稣,并寻求上天堂的保证,你可以根据基督教信仰的核心原则遵循以下步骤:
认识到自己是需要上帝宽恕的罪人。罪包括亵渎、骄傲、贪婪、情欲、忿怒、拜偶像、奸淫、偷窃、撒谎、欺骗、仇恨、赌博、酗酒、吸毒等等,没有人可以幸免。这些罪破坏了我们与神的关系,在我们与神之间制造了隔阂。圣经说:
“因为世人都犯了罪,亏缺了神的荣耀;”(罗马书3:23)。
坚信耶稣基督是上帝之子,他为你的罪孽而死并复活。
“神愛世人,甚至將他的獨生子賜給他們,叫一切信他的,不至滅亡,反得永生。”(约翰福音3:16)
向上帝忏悔你的罪过,并改过自新。
“我们若认自己的罪,神是信实的,是公义的,必要赦免我们的罪,洗净我们一切的不义。”(约翰一书1:9)
邀请耶稣进入你的生命,成为你的救主和耶和华。这意味着相信祂会拯救你,并承诺跟随祂。
“凡接待他的,就是信他名的人,他就赐他们权柄,作神的儿女。”(约翰福音1:12)
这里有一个简单的祷告,你可以用它来表达你对耶稣的信仰和承诺:
“我来到您的面前,承认我的罪孽,需要您的恩典。我相信耶稣为我的罪而死,并复活给了我新的生命。我接受他为我的耶和华和救主,将我的心和生命交给您。请原谅我,洁净我,用你的灵引导我。帮助我忠实地生活,在您的爱和目标中前行。感谢您的怜悯和救赎。奉耶稣的名,阿门。”
接受耶稣之后,重要的是要在新的信仰中成长。定期阅读《圣经》、祷告,并寻找一个当地教会,在那里你可以成为一个会支持和鼓励你的信徒团体的一部分。
通过关爱他人、分享信仰、遵照耶稣的教诲生活,用行动彰显你的信仰。
“你们若有彼此相爱的心,众人因此就认出你们是我的门徒了。”(约翰福音13:35)
相信耶稣并将自己的生命交托给祂是基督教信仰的基础,也是通往天堂永生的道路。
“他们说:当信主耶稣,你和你一家都必得救。”(使徒行传16:31)
致谢
在此,我要衷心感谢大桥教会的朴環哲牧师,他仔细阅读了整个草稿,并进行了细致的修改和必要的补充。
我还要深深感谢Kim,Yong-Cheol牧师、Kim,Jong-Kug牧师、Kim,Kyoung-A传教士和Kim,Hyun‑A女士,他们通过多次有关圣经和天文学的对话,激励了本书的出版。
此外,我还要衷心感谢夏洛茨维尔BLOO‑gene韩国教会的Im,Jun-Sub博士和牧师、ArcturusTherapeutics的Choi,Kyoung-Joo博士以及韩国化学技术研究院的Park,Chi-Hoon博士阅读了我的手稿并提供了宝贵的反馈意见。
特别要感谢我的儿子塞缪尔和丹尼尔,感谢他们对图像工作的协助。
一切荣耀归于上帝!
图片来源
1.宇宙的创造
Fig. 1.1: NASA/JPL, Fig. 1.2: Hubble Heritage Team, Fig. 1.3: R. Hurt/JPL-Caltech/NASA, Fig. 1.4: Hubble/NASA/ESA, Fig. 1.5: Wikipedia/R. Powell, Fig. 1.6: Wikimedia/D. Leinweber, Fig. 1.7: NASA/CXC/M. Weiss(left), NASA/D. Berry (right), Fig. 1.8: Stellarium, Fig. 1.9: Physics Forums, Fig. 1.10: NASA/JPL-Caltech (left), A. Sarangi, 2018, SSR, 214, 63 (right), Fig. 1.11: Wikimedia/ALMA (ESO/NAOJ/NRAO) (left), T. Müller (HdA/MPIA)/G. Perotti (The MINDS collaboration)/M. Benisty (right), Fig. 1.12: TASA Graphic Arts, Inc., Fig. 1.14: Jon Therkildsen, Fig. 1.15: www.neot-kedumim.org.il
2. 神的杰作:地球
Fig. 2.1: R. Narasimha, Fig. 2.3: NASA, Fig. 2.4: NASA/Goddard/Aaron Kaase, Fig. 2.6: Wikimedia, Fig. 2.7: Linda Martel, Fig. 2.8: Wikimedia, Fig. 2.9: NASA/ESA/H. Weaver & E. Smith (left), NASA/HST Comet Team (right), Fig. 2.10: Wikimedia/M. Bitton, Fig. 2.11: Wikimedia/John Garrett, Fig. 2.12: UK Foreign and Commonwealth Office, Fig. 2.13: Wikipedia, Fig. 2.16: Wikipedia/G. Taylor, Fig. 2.17: NASA/Caltech
3. 创造还是进化?
Fig. 3.1: Wikipedia/Yassine Mrabet, Fig. 3.2: OpenEd/Christine Miller , Fig. 3.3: Wikipedia/LadyofHats, Fig. 3.4: Wikipedia/Messer Woland & Szczepan (left), Wikipedia/LadyofHats (right), Fig. 3.5: J.E. Duncan & S.B. Goldstein, Fig. 3.6: Wikipedia/Fiona 126, Fig. 3.7: NASA, Fig. 3.8: R. Cui, Fig. 3.9: Wikipedia/Ansgar Walk, Fig. 3.10: The Whisker Chronicles, Fig. 3.11: Encyclopedia Britanica Inc., Fig. 3.12: Wikipedia, Fig. 3.13: Wikipedia/LadyofHats, Fig. 3.14: Wikipedia/J.J. Corneveaux, Fig. 3.15: Smithsonian Institution, Fig. 3.17: NRAO/AUI/NSF (left), Wikipedia/Colby Gutierrez-Kraybill (right), Fig. 3.18: Wikipedia/MikeRun, Fig. 3.20 - Fig. 3.23: Shueisha, Inc./Obara Takuya, Fig. 3.24: Wikipedia/Pinakpani, Fig. 3.25: Wikipedia/Dicklyon, Fig. 3.26: Wikipedia/Stannered (1st img), Dicklyon (2nd img), Morn the Gom (3rd img), Eequor (4th img), Fig. 3.27: M. Kitazawa/J. Plant Res., Fig. 3.28: S.R. Rahaman, Fig. 3.30: Jill Britton (pineapple), Fig. 3.32: Wikipedia/Farry (left), Wikimedia/Ivar Leidus (right).
参考资料
1.宇宙的创造
제자원 (2002), Oxford Bible Encyclopedia, Bible Textbook Co., Genesis Chap. 1-11.
Another universe existed before Big Bang? 우주먼지의 현자타임즈, 2/24/2024, https://www.youtube.com/watch?v=RckLkaVzFe0
A Big Ring on The Sky: AAS 243rd Press conference. Alexia M. Lopez, 1/11/2024, https://www.youtube.com/watch?v=fwRJGaIcX6A
Bogdan, A., et al. (2024), “Evidence for heavy-seed origin of early supermassive black holes from a z ≈ 10 X-ray quasar”, Nature Astronomy, 8, 126.
Bonanno, A., & Fröhlich, H.-E. (2015), “A Bayesian estimation of the helioseismic solar age”, Astronomy & Astrophysics, 580, A130.
Karim, M. T., & Mamajek, E. E. (2017), “Revised geometric estimates of the North Galactic Pole and the sun's height above the Galactic mid-plane”, MNRAS, 465, 472.
Lopez, A. M., et al. (2022), “Giant Arc on the sky”, MNRAS, 516, 1557.
Lopez, A. M., Clowes, R. G., & Williger, G. M. (2024), “A Big Ring on the Sky”, JCAP, 07, 55.
Lyra, W., et al. (2023), “An Analytical Theory for the Growth from Planetesimals to Planets by Polydisperse Pebble Accretion”, The Astrophysical Journal, 946, 60.
Penrose, R. (2016), The Emperor’s New Mind, Oxford University Press, Oxford, United Kingdom.
Perotti, G., et al. (2023), “Water in the terrestrial planet-forming zone of the PDS 70 disk”, Nature, 620, 516.
Sandor, Zs., et al. (2024), “Planetesimal and planet formation in transient dust traps”, Astronomy & Astrophysics, in press.
Schiller, M., et al. (2020), “Iron isotope evidence for very rapid accretion and differentiation of the proto-earth”, Science Advances, 6, 7.
Tonelli, G. (2019), Genesis: The story of how everything began, Farrah, Straus and Giroux, New York, pp 19-44
Tryon, E. P. (1973), “Is the Universe a vacuum fluctuation”, Nature, 246, 396.
Vorobyov, E. I., et al. (2024), “Dust growth and pebble formation in the initial stages of protoplanetary disk evolution”, Astronomy & Astrophysics, 683, A202.
Yi, S., et al. (2001), “Toward Better Age Estimates for Stellar Populations: The Y2 Isochrones for Solar Mixture”, The Astrophysical Journal Supplement Series, 136, 417.
2. 神的杰作:地球
Comins, N. F. (1993), What If the Moon Didn't Exist? HarperCollins Publishers Inc., New York, NY.
Gonzalez, G. & Richards, J. W. (2004), The privileged planet: How Our Place in the Cosmos Is Designed for Discovery, Regnery Publishing, Inc.
Lineweaver, C. H., et al. (2004), “The Galactic Habitable Zone and the Age Distribution of Complex Life in the Milky Way”, Science, 303 (5654), 59.
Lüthi, D. et al. (2008), “High-resolution carbon dioxide concentration record 650000 - 800000 years before present”, Nature, 453, 379.
Narasimha, R., et al. (2023), “Making Habitable Worlds: Planets Versus Megastructures”, arXiv:2309.06562.
OpenAI. (2024), ChatGPT (4o) [Large language model], https://chatgpt.com
Ward, Peter D. & Brownlee, Donald (2000), Rare Earth: Why Complex Life is Uncommon in the Universe, Copernicus Books (Springer Verlag).
3. 创造还是进化?
Abelson, P. H. (1966), “Chemical Events on the Primitive Earth”, Proc Nat Acad Sci, 55, 1365.
Behe, M. J. (2006). Darwin's black box: The biochemical challenge to evolution. Free Press.
Behe, M. J. (2020). Darwin devolves: The new science about DNA that challenges evolution. HarperOne.
Bernhardt, H. S. (2012), “The RNA world hypothesis: the worst theory of the early evolution of life (except for all the others)”, Biology Direct, 7, Article number: 23.
Chyba, C. F., & Sagan, C. (1992), “Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: An inventory for the origins of life”. Nature, 355, 125.
Cui, R., “The transcription network in skin tanning: from p53 to microphthalmia”, https://www.abcam.com/index.html?pageconfig=resource&rid=11180&pid=10026
Dembski, W. A., & Ewert, W. (2023). The design inference: Eliminating chance through small probabilities. Discovery Institute.
Danielson, M. (2020), “Simultaneous Determination of L- and D-Amino Acids in Proteins”, Foods, 9 (3), 309.
Fabre, J.-H. (2015), The Mason -Bees (Perfect Library), CreateSpace Independent Publishing Platform.
Higgins, M. (2014), “Bear evolution 101”, The Whisker Chronicles, https://thewhiskerchronicles.com/2014/01/03/bear-evolution-101/
Kasting, J. F. (1993). "Earth's Early Atmosphere." Science, 259(5097), 920.
Maslin, M. (2016), “Forty years of linking orbits to ice ages”, Nature, 540 (7632), 208.
Miller, S. L. (1953), “A Production of Amino Acids under Possible Primitive Earth Conditions”, Science, 117, 528
Mumma, M. M., et al. (1996), “Detection of Abundant Ethane and Methane, Along with Carbon Monoxide and Water, in Comet C/1996 B2 Hyakutake: Evidence for Interstellar Origin”, Science, 272 (5266), 1310.
OpenAI. (2024), ChatGPT (4o) [Large language model], https://chatgpt.com
Park, Chi Hoon (2024), “Stop codon points to GOD”, Proceedings of the 20th Anniversary KRAID Symposium
Pinto, J. P., Gladstone, G. R., & Yung, Y. L. (1980), “Photochemical Production of Formaldehyde in Earth’s Primitive Atmosphere”, Science, 210, 183.
Pinto, O. H., et al. (2022), “A Survey of CO, CO2, and H2O in Comets and Centaurs”, Planet. Sci. J., 3, 247.
Russo, D., et al. (2016), “Emerging trends and a comet taxonomy based on the volatile chemistry measured in thirty comets with high resolution infrared spectroscopy between 1997 and 2013”, Icarus, 278, 301.
Sanjuán, R., Moya, A., & Elena, S. F. (2004), “The distribution of fitness effects caused by single-nucleotide substitutions in an RNA virus”, Proc Natl Acad Sci, 101(22), 8396.
Trail, D., et al. (2011), “The oxidation state of Hadean magmas and implications for early Earth’s atmosphere”, Nature, 480, 79.
Urey, H. C. (1952). "On the Early Chemical History of the Earth and the Origin of Life." Proc Natl Acad Sci, 38(4), 351.
Wikipedia, Mutation (Distribution of fitness effects).
Wikipedia, Visual phototransduction.
Yang, P.-K. (2016), “How does Planck’s constant influence the macroscopic world?”, Eur. J. Phys., 37, 055406.
Zahnle, K. J. (1986), “Photochemistry of methane and the formation of hydrocyanic acid (HCN) in the Earth’s early atmosphere”, J. Geophys Res, 91, 2819.
关于作者
金東燦博士拥有韩国首尔延世大学天文学学士学位和夏威夷大学天文学博士学位。完成博士学业后,他在多个机构从事天文学研究,包括美国国家航空航天局喷气推进实验室/加州理工学院、首尔国立大学和弗吉尼亚大学。
金博士的研究重点是发光红外星系(LIRG)、超亮红外星系(ULIRG)、类星体和反冲超大质量黑洞。
他隶属于美国弗吉尼亚州夏洛茨维尔的国家射电天文台。
金東燦(cyberspacedckim@gmail.com)
本书是以《DIVINE GENESIS: Exploring Creation through Astronomy and Biology》为题,在美国亚马逊出版的英文书籍的中文译本。The PDF version of this book, along with translations in multiple languages, can be downloaded from divine-genesis.org.