天文学と生物学的観点から見た
創造論
はじめに
この本は、宇宙の創造、神様の傑作である地球、そして生命の起源についての議論を通じて、創造論と進化論を探求するために書かれた。
まず、宇宙の創造を論じるために、宇宙の広大さとその層状構造について簡単に紹介した後、最新の天文学的研究を通じて明らかにされた宇宙の起源と宇宙の運命について見ていく。次に、聖書の宇宙創造の記録が天文学的観測と一致するのか、地球と太陽の創造、一部の創造論者が主張するよう地球の年齢が6千年なのか、そして微調整された宇宙についても見ていきたい。
第二に、神様の傑作、地球では、地球が宇宙でどれほど特別な惑星であるかを10の主要な事実を通して調べ、これをもとに、地球が単なる偶然の産物なのか、それとも生命体の生存のために特別に創造されたのかについて調べる。
第三に、生命体の起源に関する様々な議論を通じて、進化論と創造論の妥当性を検討しようとする。そして、生命体の環境適応についての議論を通じて、ダーウィンの理論を進化論と呼ぶべきか、環境に対する遺伝的適応論と呼ぶべきか、人間が類人猿から進化したのか、知的設計論とは何かについても調べる。さらに、粒子物理学、宇宙人の存在の可能性、動物の本能、自然界で発見される数学的原理を通して創造論を見る。
最後に、皆さんを信仰に誘いたい。現代社会は物質万能主義と数多くのメディアの影響で精神的な価値が歪曲され、人間本来の崇高さと価値ある人生の基準が曖昧になっている。このような時代の中で、「私はどこから来たのか、どこへ行くのか」という根本的な質問を投げかけてみた読者がいれば、その答えとして永遠の価値を持つ福音を紹介したい。
この本を通して、読者が生命の起源がどこから来たのか、地球上のすべての生命体がどれほど大切な存在であるかを改めて考える機会になることを願っている。そして、このすべてのものを創造し、保存される創造主に対する畏敬の念を持つきっかけになることを願っている。
1. 宇宙の創造
子供の頃、田舎や高い山でキャンプをしながら、広大な夜空に輝く無数の星を眺め、暗い空を優雅に横切る流れ星を見て感嘆した思い出があるのではないだろうか。このような経験は、しばしば私たちを宇宙に対する畏敬の念に満たさせ、様々な疑問を抱かせる。宇宙にはどれだけの星があるのだろうか?地球の向こうに生命体は存在するのだろうか?宇宙はどのように始まり、どのように終わるのか?誰がこのすべてを創造したのだろうか? など、様々な疑問を抱かせるだろう。
この章では、宇宙の起源を天文学的な視点と聖書の視点から見ていく。この二つの視点を比較することで、聖書の創世記の創造の記録が天文学的観測と一致するかどうかを調べる。 また、地球と太陽のどちらが先に創造されたのか、地球の年齢が6,000年なのか、微調整された宇宙についても見てみたい。
a. 宇宙の階層構造
宇宙の起源について議論するために、まず宇宙の階層構造を見てみよう。太陽系から始まり、銀河、外銀河、銀河団、超銀河団、超銀河団Complexへと進んでいく。
i. 太陽系
「太陽系」は、太陽と呼ばれる星(恒星)とその周りを公転する8つの惑星、火星と木星の間の小惑星帯、冥王星を含むカイパーベルト、そして最後に位置するオルト雲で構成されている。ここで、星とは核融合反応によって自ら光を発する天体を指し、惑星とは自ら光を発することはできないが、星から来る光を反射して明るく輝く天体を指す。惑星は大部分が岩石で構成された地球型惑星と大部分がガスで構成された木星型惑星に分けることができる。私たちの地球は太陽から3番目に位置する地球型惑星である。
地球から月までの距離は38万4千kmで、時速1,000kmの飛行機で行くと16日かかる。地球から太陽までの距離は約1億5千万kmで、飛行機で行くと17年かかる。天文学では、地球から太陽までの距離を1天文単位で定義する。この天文単位を使うと、一番外側に位置する惑星である海王星までは30天文単位、カイパーベルトは30-50天文単位、オルト雲まで2,000-200,000天文単位になる。
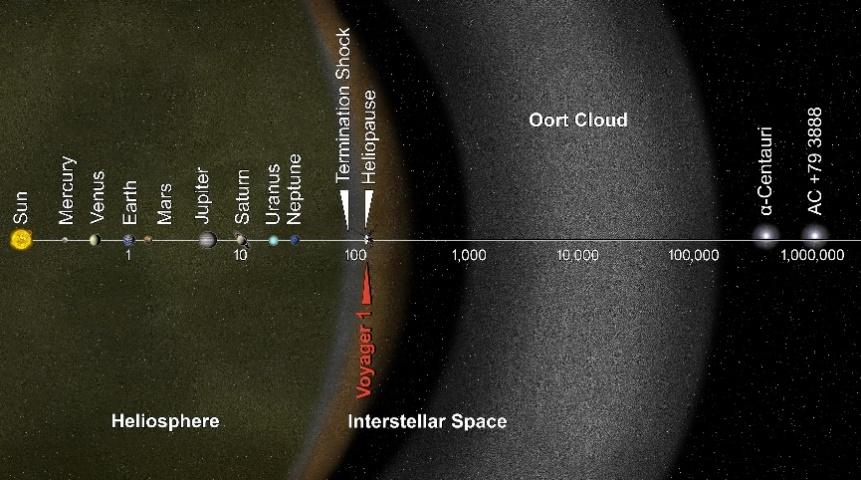
図1.1.オルトクラウドを含む太陽系
私たちが観測する彗星には短周期彗星と長周期彗星がある。短周期彗星はカイパーベルトの物体が摂動を受け、太陽の重力によって引き寄せられるものであり、長周期彗星はオルト雲の物体が摂動を受け、太陽の重力に引き寄せられるものである。このように、彗星は遠い距離から太陽を訪れるため、偏心率の大きい楕円軌道を描くことになる。
光の速度で走ると、地球から太陽までは8.3分、海王星までは4時間、太陽系の果てまで9ヶ月半、または0.79光年(1光年=光が一年間走る距離)かかる。時速1,000kmの飛行機で行くと約85万年かかる。太陽は地球の109倍の直径を持ち、質量は33万倍、自転周期は約25日である。
ii. 星の世界
オルト雲を抜けると、今度は星の世界に入る。地球から最も近い星は「プロキシマ・センタウリ」という星で、大きさは太陽の14%、質量は太陽の12%、距離は約4.2光年離れている。時速1,000kmの飛行機で行くと約457万年かかる。
私たちが夜空に見るきらめく星は、よく見ると様々な色の星があることが分かる。星の色は星の表面の温度によって変わるが、温度が低いと赤い色を、温度が高いと青白い色を帯びるようになる。鍛冶屋で鋳鉄を加熱すると、最初は赤色を帯び、温度が上がると黄色に、さらに温度が上がると青白くなるのと同じ理屈である。オリオン座のアルファ星であるベテルギウスは赤色、私たちの太陽は黄色、そして猟犬座のアルファ星であるシリウスは青白色を帯びている。このような違いをもたらす最も根本的な要因は星の質量であり、星の進化過程を決定するのも星の質量である。
星の質量は核融合の速度を決定し、これは星の明るさと寿命に影響を与える。質量が大きい星は、質量が小さい星よりも速く燃料を消費する。星が核燃料を使い果たすと、最終的には、白色矮星、中性子星、またはブラックホールに至る。星が赤色巨星の段階を経た後、中心部の質量が太陽の約1.4倍以下の星は、中心でのヘリウム核融合反応の後、ずっと収縮して白色矮星になり、その外層は膨張して状星雲を形成する。中心部の質量が太陽の約1.4倍以上、3倍以下の星は、中心部で陽子が電子と結合して中性子星になり、中心部の質量が太陽の3倍以上の星は、強力な重力により中性子星を経てブラックホールになる。中性子星とブラックホールの外層は激しく爆発して超新星となる。爆発後、その残骸が集まり、重力によって収縮することで、新しい星が再び形成される。

図1.2.様々な色を持つ星たち
現在、私たちが肉眼で見ることができる星の数は、都会ではおおよそ百個程度、田舎の奥地では約千個程度である。これらの星は、ほとんどが地球から約50光年以内に位置する非常に近い星である。
iii. 我々の銀河
私たちの銀河は棒状螺旋銀河に分類され、約2千億から4千億個の星が集まっている。銀河の大きさは直径が約10万光年、厚さが約1千光年である。太陽は銀河の中心から約2万6千光年離れており、約2億1千万年周期で銀河の中心を公転している。太陽系はペルセウス座の腕と盾座の腕の間に位置し、銀河面から約60光年上方にあり、どの方向を向いても宇宙をよく観測できる見晴らしの良い場所にある。

図1.3.私たちの銀河
iv. 銀河、銀河団、超銀河団
私たちの銀河を離れると、巨大な銀河の世界が広がる。私たちに最も近い銀河はアンドロメダ銀河で、距離は約250万光年になる。肉眼で観測が可能で、私たちの銀河に似ている。アンドロメダ銀河は毎秒約110kmの速度で私たちの銀河に接近し、約40億年後には私たちの銀河と衝突すると予想されている。銀河を視覚的な形で分類すると、渦巻銀河、楕円銀河、そして不規則銀河に分けることができる。渦巻銀河が衝突すると、明るい赤外線銀河を経て楕円銀河が作られる。

図1.4. 渦巻銀河、楕円銀河、不規則銀河
50個未満の銀河が重力的に集まっている場合は銀河群と呼び、数百から数千銀河が集まっている場合は銀河団と呼ばれる。私たちの銀河とアンドロメダを含む近くの40余りの銀河は局部銀河群に属し、局部銀河群とおとめ銀河団はおとめ超銀河団に属し、おとめ超銀河団はそれより大きなラニアケア超銀河団に属する。
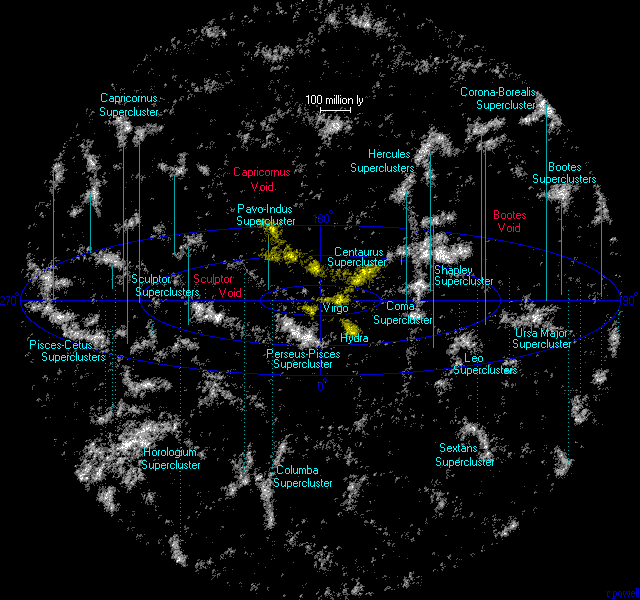
図1.5.近傍の超銀河団(黄色のラニアケア超銀河団)
超銀河団コンプレックスは、銀河フィラメントまたは超銀河団チェーンとも呼ばれ、銀河、ガス、暗黒物質の広大なネットワークで相互に接続された多数の銀河団で構成される、宇宙の巨大な大規模構造である。これらの相互接続された領域は、クモの巣のようなパターンを形成し、宇宙で知られている中で最大の構造を示している。数億光年から数十億光年に及ぶ巨大な距離に広がっており、その中でもヘラクレス・コロナ・ボレアリス万里の長城は宇宙最大の超銀河団コンプレックスである。観測可能な宇宙内で発見される銀河の数は約2兆個であり、それらを含む宇宙の大きさは約930億光年である。
b. 宇宙の創造
私たちが住んでいる宇宙はどのように始まったのだろうか?この宇宙は始まりもなく永遠前から存在していたのだろうか?それとも創造主である神様によって創造されたのだろうか? 多くの人が疑問に思うこの質問に答えるために、天文学的に明らかにされた宇宙の起源と、聖書の創世記に記録された宇宙の創造の記録を見て、この二つが一致しているかどうかを調べてみよう。
i. 天文学的に明らかにされた宇宙の創造
宇宙の起源について最も広く支持されている理論はビッグバンだ。ビッグバン説によると、私たちの宇宙は約138億年前にある特異点の爆発から始まっている。この事実は当然、一つの興味深い質問を提起する:ビッグバン以前には何が存在していたのだろうか?最新の宇宙論によると、ビッグバン以前の宇宙は何も存在しない真空状態だったという。この真空は量子揺らぎ(Quantum fluctuation)をしている状態で、ある特異点の爆発をきっかけに宇宙が始まったとされる。
ポール・ディラック以前は、真空は何もない空間として認識されていた。しかし、ディラックが相対論と量子力学を組み合わせてディラック方程式を解いたことで、真空は無ではなく、負のエネルギーを持つ粒子で満たされた空間であることが発見された。ディラックのこの発見により、物理学では真空をディラックの海と呼んでいる。ディラックの海は人間の目には静的に見えるが、決して静的ではない。むしろ、真空はハイゼンベルクの不確定性原理によって絶え間なく揺れ動いている。物質と反物質が瞬間的に「双生成(Pair creation)」され、すぐに「双消滅(Pair annihilation)」する。その時間はおよそ10-21秒となり、人間の目には見えない。もしそれを捉えるカメラがあれば、まるで狂ったように揺れ動く真空の海を見ることができるだろう。これが「量子ゆらぎ」と呼ばれる。
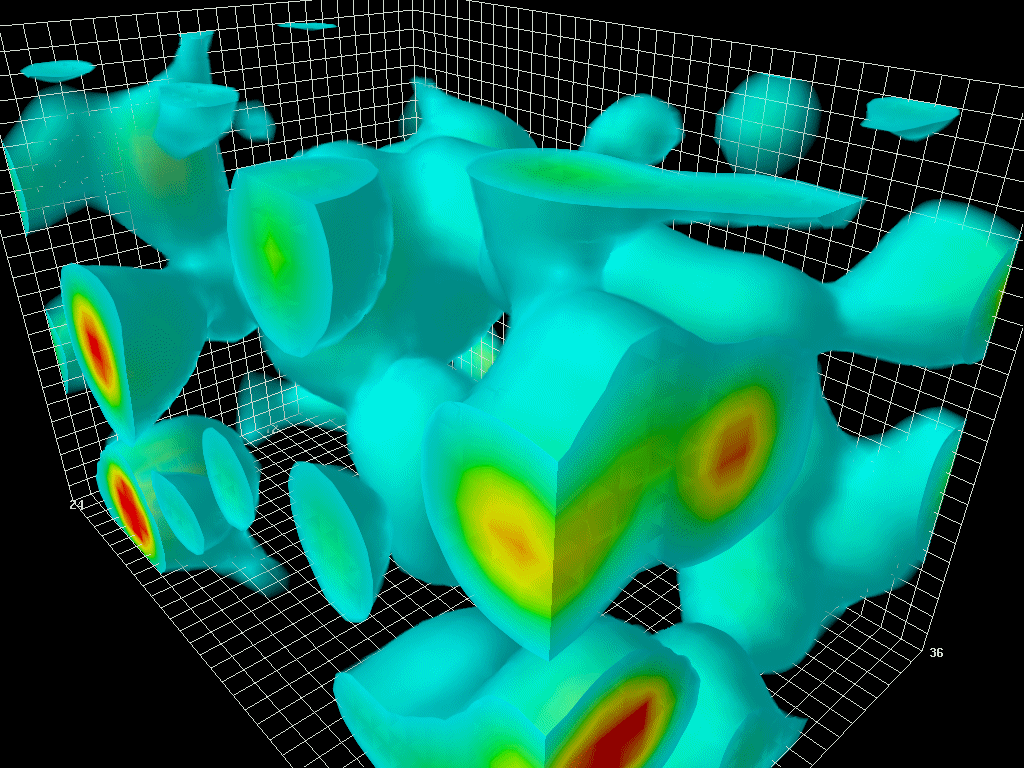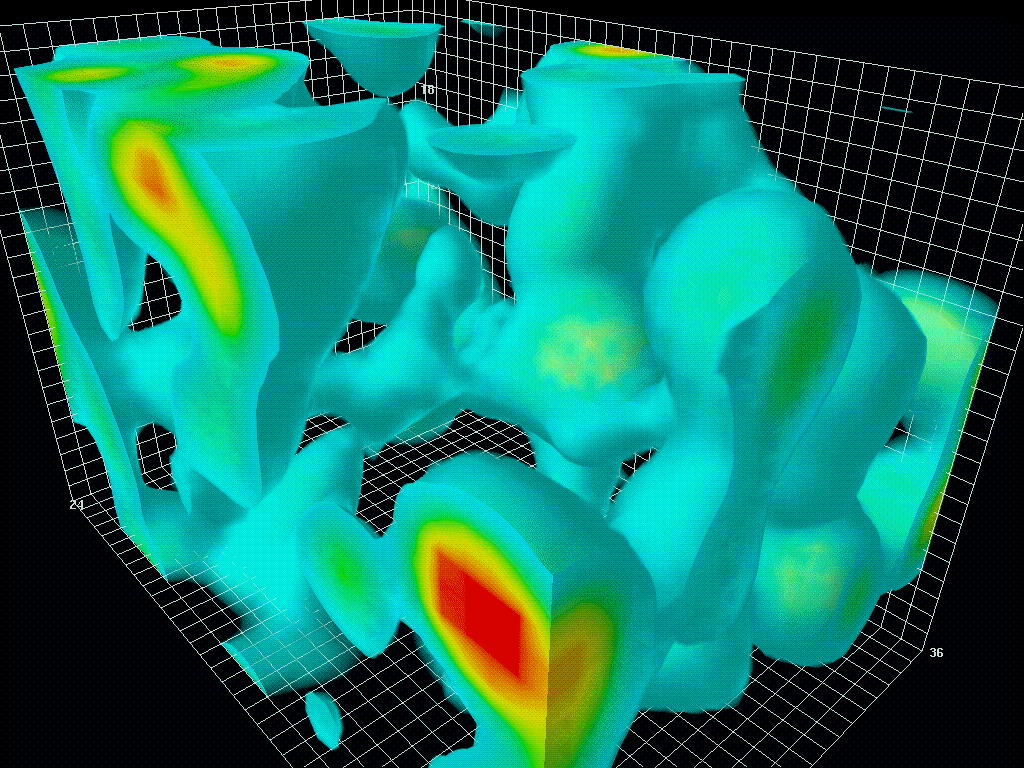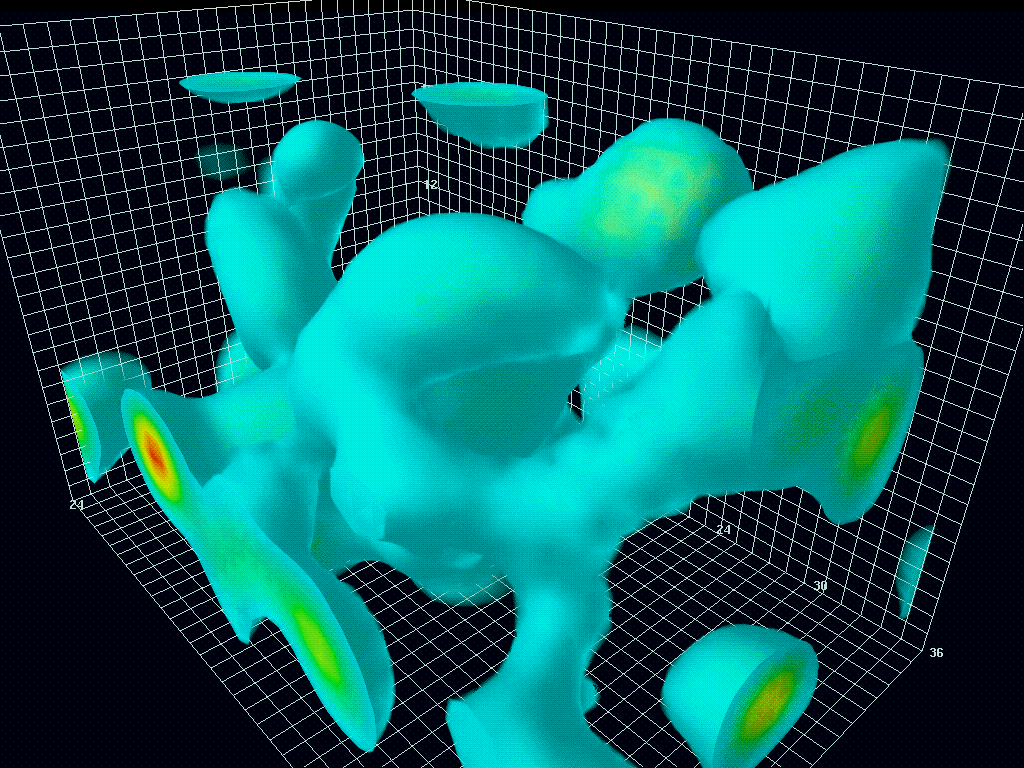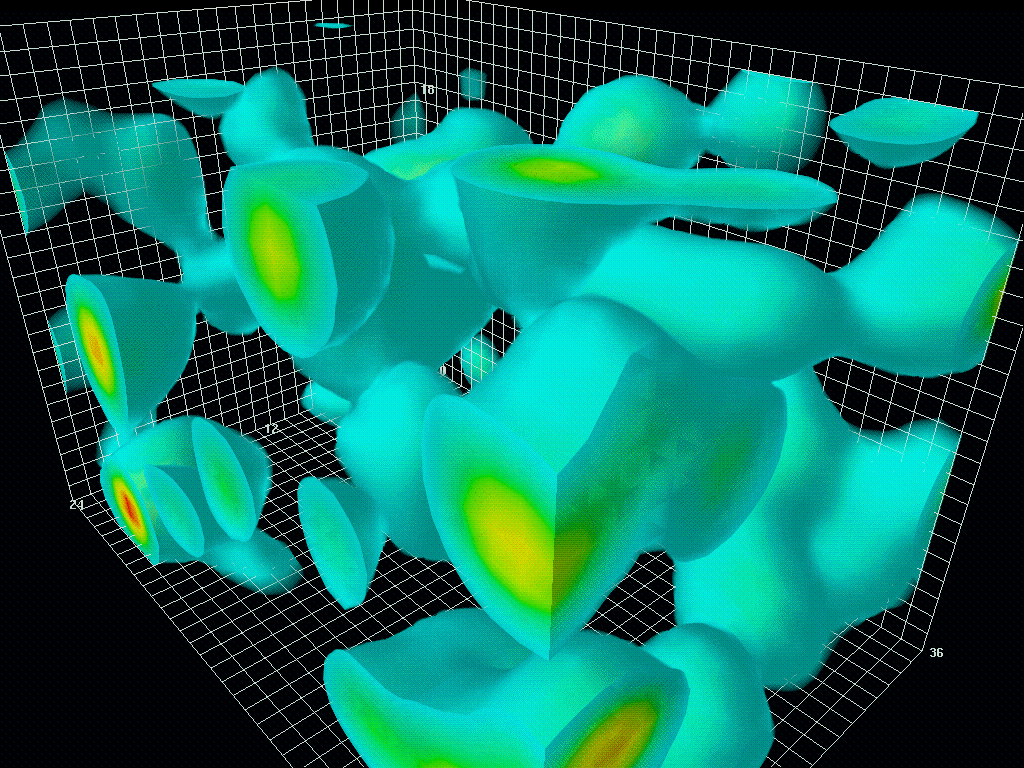
図1.6.量子揺らぎを打つ真空の3次元モデル
このように量子揺らぎが起きているある瞬間、宇宙のある特異点(Singular point)で大きな爆発が起こり、宇宙が始まった。この爆発がよく知られているビッグバン(Big Bang)だ。
ビッグバン直後の宇宙は、信じられないほどの高い温度と密度の状態にあり、非常に短い時間の間で劇的な変化を遂げる。宇宙が始まった直後のプランク時間である10-43秒から10-36秒までは、宇宙を支配するすべての力が統一された大統一時代を経る。続く10-36秒から10-32秒はインフレ時代、10-32秒から10-12秒は電荷時代、10-12秒から10-6秒はクォーク時代、10-6秒から1秒の間はハドロン時代、1秒から10秒の間はレプトン時代と呼ばれる。
レプトン時代の終わり頃、宇宙の温度が109度程度まで冷えると、非常に重要な出来事が起こる。軽粒子と半軽粒子(主に電子と陽電子)が出会い、双消滅を起こし、宇宙全体が光子(光の粒子)で満たされる。この時期を光子時代(photon epoch)と呼ぶ。光子時代は宇宙の年齢が38万年に達するまで続く。光子時代の初期、約20秒から20分の間、核合成時代を経ながら陽子、中性子、ヘリウムが生成され、物質が作られる準備をする。ビッグバン直後から光子時代までの期間を放射線支配時代と呼ぶが、時間的に見れば、その大部分、ほぼ100%は光子によって支配される光子時代と見ることができる。光子時代に生成された光子は、プラズマ状態の自由電子や陽子によって散乱され、自由に移動することができない。したがって、この時期の宇宙は、まだよく観測ができない不透明な状態で存在する。
光子時代の終わりに、宇宙の温度がある程度下がり始めると、再結合時代が始まる。この時、核合成時代に作られた自由電子と陽子が結合して中性水素が作られる。そうなると、プラズマで満たされた宇宙が徐々に透明になり、私たちが空と呼ぶ空間が現れる。そして、光子時代に生成されたが、プラズマに閉じ込められていた光子が、ようやく透明になった宇宙を歩き回り、私たちに観測され始める。このように自由に動く光子は非常に明るい光として観測され、現在の宇宙マイクロ波背景放射の起源となる。
再結合時代に物質が作られ、物質支配時代に入ると、この時に形成された物質は、現在私たちが見にしている星や銀河を生成する材料となる。星と銀河が生成された後、宇宙はビッグバンの余波によって膨張を続ける。宇宙の年齢が約98億年になった時は、目に見えない暗黒エネルギーが宇宙を支配しており、この時期は暗黒エネルギー支配時代と呼ばれる。現在の宇宙は、この暗黒エネルギーの影響により、銀河の距離が互いに遠ざかる加速膨張の状態である。
ii.宇宙の運命 (再起動 ?)
加速膨張している宇宙の運命はどうなるのか?私たちの宇宙の未来は、宇宙全体の密度に依存している。宇宙の密度が臨界密度(10-29g/cm3)を上回る場合(閉じた宇宙)、宇宙は膨張を続けるが、ある時点で膨張を止めて再び収縮することになる。これをビッグクランチ(Big Crunch)と呼ぶ。逆に、密度が臨界密度を下回る場合(開放宇宙)、宇宙は加速膨張を続け、最終的にはビッグリップ(Big Rip)と呼ばれるシナリオにつながる。この場合、膨張が進むにつれて、宇宙の温度は徐々に下がり、星の生成に必要な星間媒体が枯渇し、新しい星が生まれなくなる。そして、既存の星々も燃料を使い果たし、やがて光を発せなくなる。
その後、宇宙の年齢が約1032年になると、大統一理論の予測通り、陽子崩壊が始まり、物質は徐々に消えていく。さらに約1043年が経過すると、ブラックホールさえもホーキング輻射によって蒸発し始める。すべての粒子が崩壊し、ブラックホールが蒸発すると、宇宙は放射に満ちた暗黒の時期へと突入することになる。宇宙の温度は絶対零度まで冷却され、最終的にはすべてが暗く空虚な状態になり、ビッグバン以前の量子揺らぎを経験している宇宙の状態に似てくる。
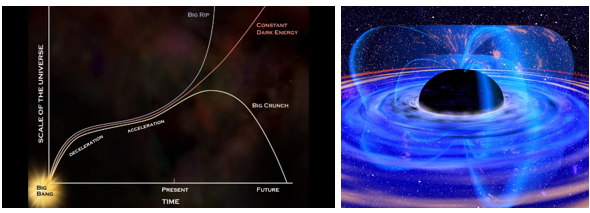
図1.7.宇宙の運命と蒸発するブラックホール
宇宙の運命についてのもう一つの理論は、宇宙が循環しているという循環宇宙論である。最近、地球から北斗七星の方向に70億光年離れた宇宙において、二つの宇宙巨大構造が発見された。2022年に発見されたGiant Arcと、2024年に発見されたBig Ringである。これらの巨大構造は、宇宙原理の均質性に反する可能性があり、新たな何らかの適切な説明が必要になる。考えられる仮説の一つとして、宇宙の紐(Cosmic string)または、以前のビッグバンによって生まれた超巨大ブラックホールの残骸である可能性が挙げられている。実際、ホーキング輻射によるブラックホールの蒸発速度は極めて遅い。太陽質量程度のブラックホールの蒸発時間は約1067年、超巨大ブラックホールの蒸発時間は約10100年かかる。しかし、これらのブラックホールが出会い、衝突を起こしたとすれば、非常に強力な重力波が発生する。そして、今回発見されたビッグリングが、かつての宇宙で発生した重力波の痕跡(Hawking Point)である可能性があるということになる。
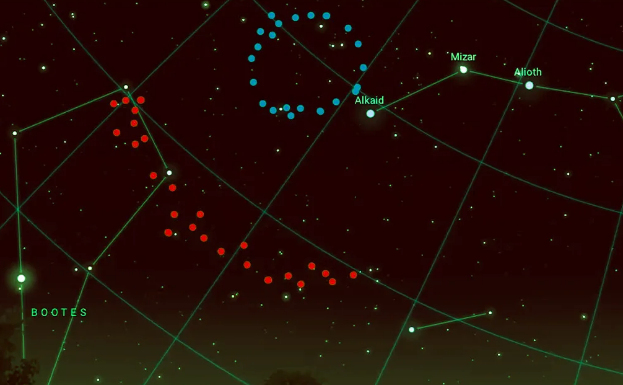
図1.8.ビッグリング(青)とビッグアーク(赤)
この解釈は、ロジャー・ペンローズの等角循環宇宙論(CCC)と関連している。CCCは一般相対性理論に基づいた宇宙論的モデルで、宇宙が膨張を続け、すべての物質が消滅し、ブラックホールだけが残された状態になると、再び新しいビッグバンが始まるという仮説である。個人的にCCCが興味深い理由は、銀河進化に関するいくつかの問題を解決する可能性を示唆しているからである。ブラックホールの質量と恒星の速度分散の間には相関関係が存在する事が明らかになった。ブラックホールの質量は一般的に銀河質量の約0.1%程度に相当するという。しかし、宇宙初期のブラックホールの質量を見ると、この比率が合わない。例えば、ビッグバンから約5億年後の銀河UHZ1のブラックホールは、銀河全体の質量よりも大きい。これを説明するために様々な仮説が提示されたが、CCCを適用すれば簡単に解決できる。つまり、以前の宇宙で生成されたブラックホールが新しい宇宙の「種子ブラックホール」になり、周囲のガスや塵を集め、それによって銀河が形成されたと仮定すれば、初期宇宙に観測される異常に大きなブラックホール質量の疑問を効果的に説明することができる。

図1.9.等角循環宇宙論
では、膨張する宇宙の中で再び新しい宇宙が生まれるというのはどういうことだろうか。3次元空間では、ある物質が散らばるということは、一点に集まることの反対の言葉である。しかし、4次元空間から見た3次元の散らばりは、4次元空間の位相構造によって一つの特異点に収束する構造になっている可能性もある。例えば、クラインの壺の内部表面から出てきた物体が入口では一つの点に収束して入るように、3次元での膨張が4次元では収束する形で現れるかもしれない。もしそうであれば、等角循環宇宙論は自然に説明できる。もしかすると、4次元空間に住む小学生なら、4次元から見た3次元の散乱は、4次元で一つの特異点に集まることができることを当たり前のように理解しているかもしれない。
この場合、限りなく膨張する宇宙は、やがて巨大な真空の海に変わり、量子揺らぎが起こりながら、次の宇宙の誕生が準備される。このように膨張する宇宙がある特異点へと収束していくと、その特異点の真空エネルギー密度がどんどん上がり、ある時点ではすべてのエネルギーを保持できず、大爆発が起こる。これが次の宇宙のビッグバンかもしれないという話だ。ビッグバンの後も、特異点に真空エネルギーが引き続き集まってくると、それが暗黒エネルギーの起源となり、新しくできた宇宙膨張の動力源になることもあるだろう。
iii.キリスト教の聖書に記された宇宙の創造の物語
キリスト教の経典である聖書は、神の言葉が記録された本であり、歴史上最も多く読まれているベストセラーである。聖書の最初の本である創世記には、天地万物の創造についての記録が記されている。この章では、創世記に記録された宇宙の創造が天文学的な事実とどのように一致しているのかを調べてみよう。
a) 神の創造宣言
創世記1章1節に次のように記されている。
「初めに、神が天と地を創造した。」(創世記1:1)
上記の箇所で見られるように、聖書は神様が直接宇宙を 創造したと具体的に記録している。続いて宇宙の創造についての説明が続くが、創世記1章2節を見ると、宇宙が始まる前はどのような状態であったかを描写している。
「地は形がなく、何もなかった。やみが大いなる水の上にあり、神の霊は水の上を動いていた。」 - 新改訳聖書
「地は混沌であって、闇が深淵の面にあり、神の霊が水の面を動いていた。」 - 新共同訳聖書
ここで「地」は、神が後に形成される物質、すなわちバリオン物質を意味する。そして「形がなく」は英語聖書では「without form」と訳されている。したがって、「地は形がなく」という表現は、初めにどんな物質も形成されなかったことを意味する。次に、'何もなかった'は英語聖書で「'void」と訳され、何も存在しない真空状態を意味する。 だから、この二つの翻訳を合わせると「地は形がなく、何もなかった」は、太初に何もない真空状態が存在したことを意味する。ここまでの解釈に無理はないと思う。
しかし、その次の文「闇が深淵の面にあり」(新共同訳聖書)が深遠な意味を含んでいる。「闇」はヘブライ語で「וחש」であり、これは光が創造される前の根本的な暗闇の状態を意味する。続く言葉である「深淵」が非常に核心的な意味を持つ。深淵のヘブライ語は「תהום」で、'揺れる'または'沸騰する'という意味を持つ「הום」に由来する言葉で、深さの分からない海または空間が揺れる状態を表す。
これを先の文章と結びつけると、「何もない暗黒状態の真空が揺れ動く」という意味になる。この解釈は、先に述べた天文学的事実、つまりビッグバンが始まる前の「暗黒状態の真空が量子揺れを打っている状態」を正確に表現している。
b) 創造初日の光の創造
真空の量子揺れを記録した創世記1章2節に続き、創世記1章3節には、神が創造の初日に行った驚くべき出来事が記録されている。
「そのとき、神が『光よ。あれ。』と仰せられた。すると光ができた。」(創世記1:3)
この箇所は、神様が宇宙を創造する時、最初に光を創造されたと記述している。この創世記に記録された光の創造は、ビッグバン直後の光子時代に光が生成され、宇宙を満たしてきた天文学的事実と正確に一致する。光子の時代に生成された光は、宇宙が透明になり、空が形成されたとき、私たちに観測されるようになった。
c) 創造二日目の天の創造
創世記の創造の二日目の主な出来事は、天の創造だ。
「こうして神は, 大空を 造り、...、神は、その大空を天と名づけられた。... 。」( 創世記1:7, 8)
創世記に記された天の創造は、ビッグバンによって発生した光子時代の後に来る再結合時代と関連している。光子時代の宇宙は、密度が高く熱いプラズマで満たされた状態だった。このプラズマは光子を散乱させ、光子の自由な移動を妨げ、宇宙を不透明な状態にしたため、私たちが空と呼べる空間は存在しなかった。しかし、再結合時代になって、宇宙の温度が徐々に下がり、電子と陽子が結合して中性水素を形成した。こうなると、プラズマに閉じ込められていた光子が自由に移動できるようになり、その結果、私たちが空と呼ぶ空間が現れるようになる。したがって、創世記の創造第二日の天の創造は、再結合の時代に天が現れた出来事を正確に描写している。
次の表は、聖書に記録された宇宙の創造と天文学的に明らかにされた宇宙の生成過程をまとめたものである。この表は、創世記の創造の記録が天文学的事実と驚くほど正確に一致していることを示し、神がこれらの事実を科学が明らかにするはるか以前から、すでに聖書を通じて教えてくださったことを確認させてくれます。
聖書の創世記 | 天文学 |
真空の量子揺らぎ (創造以前 - 創世記1:2) | 真空の量子揺らぎ (ビッグバン以前) |
光の創造 (創造の初日 - 創世記1:3) | 光の創造 (光子時代) |
天の創造 (創造の第二日 - 創世記1:7-8) | 天の創造 (再結合時代) |
表1.1.聖書の創世記と天文学の宇宙創造の比較
c. 地球と太陽、どちらが先に創造されたのか?
創世記では、三日目に神様が水と共に地球を創造し、四日目に地球を照らす光明である太陽を作ったと記録されている。この記録は、地球が太陽より先に創造されたことを示しているが、果たしてこれが天文学的に本当なのか調べてみよう。
星や惑星は星間分子雲(molecular clouds)から形成される。分子雲の構成成分は約98%がガス(水素約70%、ヘリウム約28%)であり、残りの2%は塵(炭素、窒素、酸素、鉄など)である。星と木星型惑星の大部分はガスでできており、地球型惑星の大部分は塵で形成されている。
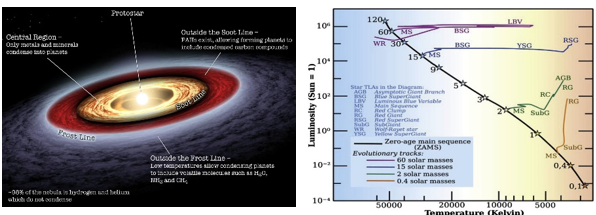
図1.10.原始星と原始惑星系円盤、そしてH-R線
分子雲の中心で重力収縮が起こると、原始星(protostar)が生成される。同時に、原始星の周りには惑星が形成される揺りかごである原始惑星系円盤(protoplanetary disk)が形成される。原始星の内部の温度はまだ核融合を起こすほど高くなく、重力収縮による衝撃波で温度が多少上昇するが、可視光線では観測されない。原始星が引き続きゆっくりと収縮すると、前主系列星(pre-main sequence star)に進化し、林トラック(Hayashi track)とヘニエイトラック(Henyey track)という星の進化過程を経る。この段階を過ぎて原始星内部の温度は核融合が可能な1千万から2千万度まで上昇し、収縮が止まると私たちが夜空で見ることができる光明体、つまり真の星が誕生する。この時から星の一生は主系列星(main sequence star)の段階に入り、核融合が始まる時点をゼロ年齢主系列(zero age main sequence)と呼ぶ。恒星進化理論と太陽地震学(helioseismology)研究によると、太陽は約4千万から5千万年の間、主系列星の状態を経た後、零点主系列星に到達して核融合を開始し、自ら光を発する光体となる。
惑星の場合、原始惑星系円盤を構成するガスと塵が衝突し、砂の大きさ、砂利の大きさ、岩の大きさ、数十または数百キロメートルサイズの小惑星(planetesimal)、そしてこれらの小惑星が集まった原始惑星(protoplanet)が作られ、最終的に惑星に進化する。まとめると、地球と太陽は同じ分子雲から始まったが、地球は原始惑星系円盤から原始星を経て誕生し、太陽は主系列星を経て主系列星として誕生する。
原始惑星系円盤の塵の雲から惑星が生成される過程は最近になって活発に研究が進んでいる。最近発表された論文によると、原始惑星系円盤の塵の雲から地球型惑星である水星、金星、地球、火星の質量を合わせた質量の砂利が、中心の主系列星から4天文単位内に形成されるのにかかる期間は約30万年であるという。また、1ミリメートルサイズの砂から原始星を経て地球サイズの惑星が誕生する期間は、約数百万 年かかると予測されている。
これらの惑星生成理論は、観測によって確認することができる。次の画像は、世界最大のサブミリメートル望遠鏡であるALMAで観測したおうし座HL Tau(左)とセンタウルス座PDS 70(右)である。HL Tauの質量は太陽の約2倍、年齢は約100万年程度で、中心には主系列星が形成されている。しかし、このように星が形成される初期段階にすでに複数の惑星が作られ、その周りを回っていることを示している。主系列星の周りの円盤を見ると、複数のギャップ(gap)が見られ、これはまさに惑星が形成され、周囲の雲を一掃した跡である。
PDS 70の質量は太陽の約0.76倍で、年齢は約540万年であり、中心には主系列星が成長している。PDS 70の内側の軌道を見ると、2つの木星型惑星PDS 70bとPDS 70cが形成され、公転していることが観測されており、HL Tauで見られた塵の雲がこの2つの惑星によってほとんど除去されたことを示している。2023年、ジェームズ・ウェブ宇宙望遠鏡で中心部分を分光観測したところ、図のスペクトルに示すように多量の水が観測され、観測された領域は地球型惑星が生成される場所であり、その中に2つ以上の地球型惑星が形成されていることを示唆された。
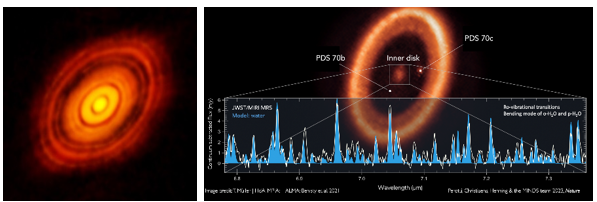
図1.11.HL TauとPDS 70
ここで重要な事実は、原始惑星系円盤の塵の雲から約540万年後、水とともに地球型惑星が生成されたという点であり、これは創世記の創造第三日に水と地球が一緒に創造されたという記録と一致する。この時までにかかった時間が540万年であり、長く見積もて1000万年かかったとしても、この期間は太陽が星になるまでの期間である4千万年から5千万年よりもはるかに短いことを示している。
したがって、創世記に記録された地球が先に創造され、次に太陽が創造されたという記録が天文観測を通じて事実であることが証明される。
神様が三日目に行われたもう一つの主要な出来事は、植物と木の創造である。無神論者や進化論者はしばしば「地球が先に創造され、その後に太陽が創造されたなら、地球と一緒に創造された植物は太陽がないのにどうやって生き残ったのか」という疑問を提起する。しかし、この問題は星の進化過程を知れば簡単に解決される。重力収縮によって形成されるT Tauri星の表面温度は約4,000から5,000度程度になり、黒体放射のピークは可視光線領域に位置する。また、主系列星であるT Tauri星段階の太陽は、現在の主系列星である太陽よりも数倍ほど大きく、地球上の植物が光合成をするのに十分なエネルギーを供給することができた。したがって、太陽が主系列星に到達する前でも植物の生存には問題はなかった。
d. 地球の年齢は6,000年か?
若い地球創造論者は聖書の創世記の創造記録を被造物の立場で解釈し、地球と宇宙の年齢が6,000年程度であると主張する。しかし、地質学、天文学、物理学など様々な分野の様々な科学的証拠によると、地球の年齢は約46億年、宇宙の年齢は約138億年となる。このような科学的事実にもかかわらず、若い地球創造論者たちは依然として地球の年齢が6,000年であると主張する。本題に入る前に、まず、地球と宇宙の年齢が少なくとも数百万年であるという事実を簡単に理解できるよう、いくつかの例を見てみよう。
地球の年齢が少なくとも数百万年であることは、プレートテクトニクスを認めればよい。地球の地殻は複数のプレートで構成されており、これらのプレートの動きによって地震が起こる。この事実を否定する人はいないだろう。熱点(hot spot)は、地殻の下にあるマントル深部からマグマが流れ出る地点で、その中心部が固定されている。マグマが地殻の上に噴出し、冷えると地面が作られるが、その代表的な例がハワイ諸島である。ハワイ本島であるビッグ・アイランドのキラウエア島では現在も火山活動が活発に行われており、噴出したマグマが海水に冷やされながら地面が作られている。こうして形成された土地は、プレートテクトニクスによって北西に年間約7~10cmずつ移動し、このプロセスが繰り返され、ハワイ諸島の島々が形成された。これは現在も観測できる紛れもない事実だ。
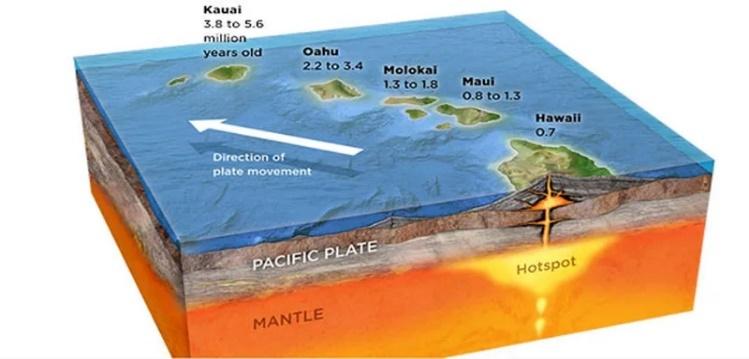
図1.12.火山活動によって形成されたハワイ諸島
地殻板の移動速度を考慮したハワイ諸島の年齢は、北西方向にビッグ・アイランドは40万年、マウイ島は100万年、モロカイ島は150万年から200万年、ワイキキのあるオアフ島は300万年から400万年、カウアイ島は5百万年ほどである。ビッグ・アイランドの場合、土地の大部分がまだ黒い火山土で覆われており、風化がほとんど行われていないことがわかる。一方、カウアイ島は比較的風化が進み、樹木がよく育っているため、庭園の島と呼ばれている。これらの事実は、地球の年齢が少なくとも数百万年以上であることを直接的に証明している。
宇宙の年齢が少なくとも数百万年以上であることは、光が1秒間に30万kmを移動することを認めれば理解できる。この事実も否定できない。太陽は地球から1億5千万km離れている。私たちが今、暖かい太陽光を受けているということは、その光が太陽から8.3分前に生成された光であることを意味する。太陽は月の約400倍ほど大きいが、遠く離れているため、視直径は月と同じように見える。アンドロメダ銀河は私たちの銀河と大きさが似ているが、250万光年離れているため、月の4倍ほど小さく見える。私たちがアンドロメダ銀河を見ることができるということは、その光が250万光年前にアンドロメダで生成され、私たちに届いたことを意味する。アンドロメダ銀河を見た人は、この事実を否定することはできないだろう。この事実は、宇宙が少なくとも数百万年であることを直接的に示唆する証拠となる。
このような事実にもかかわらず、依然として地球の年齢が6,000年であると主張することは、福音を広めるための助けになるどころか、障害となり、潜在的に多くの人々を福音から遠ざける可能性がある。したがって、無理に若い地球創造論を主張するよりも、聖書の創世記を注意深く読んで解決策を探す方がより合理的であろう。
人間にとって時間は物理量であり、常に現在から未来へ流れるものとして認識されてきた。しかし、 この時間に対する固定観念を取り除き、この問題にアプローチすれば、何らかの解決策が生まれるかもしれない。この事実を念頭に置いて、さらに詳しく調べてみよう。
i. 創世記の創造の6日間
まず、創世記の記録をもとに、宇宙の年齢を推定してみよう。創世記によると、神様は6日間かけて宇宙とその中のすべてのものを創造された。神様が最初に創造された人間であるアダムは何歳まで生き、その子孫は何歳まで生きたのか、その系譜が聖書の創世記5章に詳しく記録されている。
「これは、アダムの歴史の記録である。... ノアが五百歳になったとき、ノアはセム、ハム、ヤペテを生んだ。」(創世記5:1-32)
アダムから始まり、彼の子孫であるノアの時代の洪水までの期間を聖書に記録されている通り合わせると1,656年になる。ノアの洪水の時期については様々な意見があるが、様々な資料を参考にすると、ノアの洪水は今から約2,400年前に発生したと推定される。したがって、アダムが創造された後、ノアの洪水までが1,656年であり、ノアの洪水の時から今まで約4,400年が過ぎたので、聖書に言及された宇宙の年齢は創造7日+1,656年+ 4,400年で約6,056年になる。これが、地球の年齢が6,000年であると主張する若い地球創造論者たちの根拠となる。
この問題を解決するために、もう一度創世記を注意深く見てみよう。アダム以降の記録は正確なものと推定され、ノアの洪水が起きた時期が今から4,400年前であろうと44,000年前であろうと、私たちの宇宙の年齢138億年には何の影響も及ぼさない。では、創世記に示された宇宙の年齢が間違っているのだろうか、それとも創世記の記録の中にこの問題を解く鍵が含まれているのだろうか?おそらく創世記1章を注意深く読み、少しの天文学的知識を持っている人であれば、創世記の宇宙創造の最初の7日間にその鍵が含まれていることに気づくことだろう。
その推論は次の通りである。被造物の立場で定義する一日は、被造物が住む惑星の自転速度によって定義される。人間が金星に創造された場合、1日は243日であり、木星に創造された場合、1日は10時間である。人間が地球に創造されたため、私たちの1日は、地球が1回自転するのにかかる時間、すなわち24時間である。また、人間の観点から一日を定義するためには、まず地球があり、地球を照らす太陽がなければならない。ところで、地球は創造の三日目に、そして太陽は創造の四日目に創造された。しかし、人間の一日が定義される前に、神様は一日という言葉を使われた。したがって、創世記の創造7日間の一日は、私たち人間が定義する一日ではなく、神様の視点で定義された一日だと見ることができる。若い地球創造論者たちが創世記の一日を人間の一日とみなし、地球の年齢を6千年と主張した誤りがここで生じ、実際には地球の年齢の秘密はこの創造7日間の時間の中に含まれている。
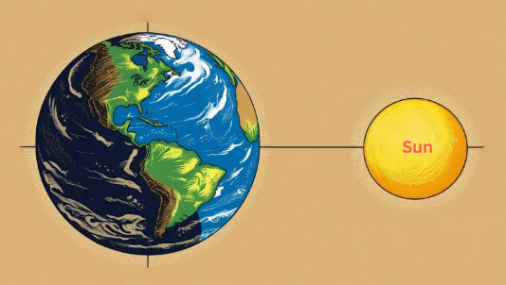
図1.13. 地球と太陽がなければ、一日を定義することができない。
では、創世記の創造の7日間の時間は、人間の時間ではどのくらいなのだろうか?正確な答えは分からないが、創世記に記述された創造の出来事とビッグバンの出来事を比較すれば、おおよその時間を推定することができるだろう。
創世記の創造第一日の代表的な出来事は光の創造である。天文学的には、ビッグバン直後の光子時代がこの出来事に該当するため、創造の初日の時間は38万年となる。創造第二日の代表的な出来事は天の創造である。光子時代以降の再結合時代がこの出来事に該当するため、創造第二日の時間は10万年である。創造三日目の代表的な出来事は地球の創造である。 前に見たように地球型惑星が生成されるのに約1000万年かかるので、創造第三日の時間は約1,000万年より大きい。 創造第四日の代表的な事件は太陽の創造だ。太陽が形成される時間は4-5000万年かかるので、創造4日目の時間は4000万年以上ですだろう。 これらの結果をまとめると次の表のようになる。
創造 | 創世記事件 | 天文学イベント | 時間 |
初日 | 光の創造 | 光子時代の光の創造 | 380,000年 |
二日目 | 天の創造 | 再結合時代の天の創造 | 100,000年 |
三日目 | 地球の創造 | 地球の創造 | >1,000万年 |
四日目 | 太陽の創造 | 太陽の創造 | >4,000万年 |
表1.2.創世記の創造時間
このように、創造の四日目まで見てみても、私たちは驚異的な事実を発見する。神様の視点から見た創造の一日は、人間が定義した24時間に比べて非常に長い時間であり、その中には数十万年という短い時間もあれば、数千万年以上に長いこともあるなど変化するということだ。これをどのように理解すればよいだろうか?おそらくある観点からは、これが驚異的な結果ではなく、すでに予想されていた結果だと見ることができる。
ii. 時間の創造者
創世記で使われている「日」のヘブライ語はヨム(יום)である。ヨムはいくつかの意味に解釈されることができるが、その中には期間が決まっていない時間を表す意味もある。創造主である神にとっての時間は、時間の限界の中で生きる人間とは全く異なる概念である。なぜなら、神は時間を創造し、支配しておられるからだ。少し聞き慣れない言葉かもしれないが、聖書には神様が時間を支配する出来事が何度も登場する。
まず、新約聖書のペテロ後書を見ると、次のような聖書箇所がある。
「しかし、愛する人たち。あなたがたは、この一事を見落としてはいけません。すなわち、主の御前では、一日は千年のようであり、千年は一日のようです。」(ペテロ後書3:8)
この聖句は、神の約束を待つ者たちが焦らずに忍耐をもって待つことを勧める意味もあるが、神の時間概念は人間の時間概念とは全く異なり、神は時間を増やしたり減らしたりすることができる方であることを示唆する聖句だと思う。
私たちは、時間が固定された物理量ではないことを知っている。特殊相対性理論によれば、動いている観測者の時間は、同じ慣性フレームで静止している観測者よりもゆっくり流れる()。また、一般相対性理論では、強い重力場下では時間がゆっくり流れることがわかっている()。このように、時間の伸びは相対論で説明できるが、時間の短縮については現在の人間の科学知識では理解しにくい部分である。
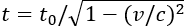
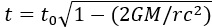

図1.14.時間膨張を示す図
ところが、聖書には、神様が時間を自由に増やしたり減らしたりするだけでなく、さらには時間を止めることもできることを示唆する出来事が記録されている。旧約聖書のヨシュア書を見ると、次のような箇所がある。
「日は 天のまなかにとどまって, まる 一日ほど 出て 來ることを 急がなかった。 」(ヨシュア10:13)
この箇所は、ヨシュアがアモリ人と戦っていた時、彼の祈りに対する神の答えとして、ほぼ一日間、太陽が中天にとどまった出来事を記録している。
しかし、これよりさらに驚くべき出来事が旧約聖書列王記第二に登場する。それは、神様が重病で死の危機に瀕したヒゼキヤ王の切実な祈りを聞き、彼に15年の命をさらに許可されたしるしとして示された時間の逆転現象である。
「預言者イザヤが主に祈ると、主はアハズの日時計におりた日時計の影を十度あとに戻された。」(列王記第二20:11)
ここで言及された日時計はアハズ王の時代に作られたもので、神様がこの日時計の時間を10段階過去に戻したということである。この時間の逆転現象は、時間の停止や短縮よりも理解するのが難しい事実である。しかし、時間を創造し、その時間を速く、遅く、または停止させることができる神様が、時間を逆さまに流すことも、まるでゲーム機のジョイスティックを逆に回すように、簡単にできることだろう。

図1.15.アハズの日時計モデル
e. 微調整された宇宙
オーケストラの演奏会に行くと、演奏が始まる前に、すべての団員が自分の演奏する楽器を調律しているのを見ることができる。この時、オーボエの「ラ(A)」の音を基準に全ての楽器が調律される。これは、オーボエが他の楽器よりも高音でもバランスの取れた音色を持ち、比較的安定した音を出すからだ。 また、「ラ(A)」の音を基準にするのは、国際的に標準化された調律基準が440Hz( )に設定されており、この音が人の耳に最もよく聞こえる音色だからだという。こうして調律が終わると、指揮者が登場し、指定された曲を演奏する。もし楽器が調律されていない状態で演奏が始まったらどうなるだろうか?おそらく文字通り不協和音の演奏会になるだろう。
微調整された宇宙(Fine-tuned universe)とは、言葉の意味通り、宇宙を構成して運行する様々な物理定数が宇宙の維持と生命体の生存のために極めて精密に調整されているという事実を意味する。
前述したように、ビッグバンで宇宙が生成された時、宇宙の質量密度が臨界質量密度より大きかった場合、宇宙は生成されると同時に収縮し、逆に臨界質量密度より小さかった場合、急速に膨張し、その結果、星や銀河、そして生命体が誕生することができなかっただろう。しかし、私たちの宇宙の質量密度は臨界質量密度とほぼ一致するように絶妙に調整され、現在の宇宙が形成され、維持されることができた。
ペンローズは彼の本《The Emperor's New Mind》(pp 440-447)で、ブラックホールエントロピーに対するバーケンシュタイン-ホーキング式を使用して、ビッグバン直後の宇宙の質量密度が臨界質量密度に似ている確率を計算したが、その確率は10の10123指数乗数の1であった。この確率は、全世界の海辺にある砂の中からある特定の砂を選ぶ確率よりもはるかに低く、決して自然界では起こり得ない確率である。
さて、そうして誕生した宇宙が順調に稼働している背後には、もう一人の重要な主人公がいる。彼らはまさに物理学の基本的な定数であり、その値は生命体の存在にとって絶妙に設定されている。宇宙を支えている代表的な物理定数は、重力定数、光の速度、プランク定数、ボルツマン定数、誘電率(vacuum permittivity)、微細構造定数(fine-structure constant)などがある。しかし、これらの定数の値が現在のものと異なっていたら、私たちはこの世に存在できなかっただろう。
例えば、重力定数が今より小さかったら、星や銀河の形成が不可能だっただろうし、今より大きかったら、私たちが地球のような惑星に住むこともできなかっただろう。プランク定数の場合、現在の値より大きかったら、地球に到達する太陽のエネルギーが減少し、深刻な気候変動が起こっただろう。また、物質の結合力が弱まり、容易に破壊され、光合成効率が低下し、植物の生存に影響を与えただろう。
ところで、これらの物理定数の中で、物理学者の関心を最も多く集めた定数が、次に示す微細構造定数αである。
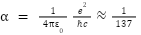
この定数は、まるで薬屋の甘草のように、様々なところで頻繁に登場する特別な定数で、他の定数と違って次元を持たず、その値は約1/137となる。この定数は、電子と陽子、あるいは二つの電子間に働く電磁力の強さを示す。
もしこの定数の値が1/137より大きければ、原子間の相互作用がより強くなり、原子の大きさが小さくなり、化学反応が容易になる一方、水素のような軽い元素は形成されにくくなる。水素は、ほぼすべての星の構成要素であり、核融合の原料であるため、水素の形成が不可能であれば、星だけでなく、その星をエネルギー源とする生命体の生存も不可能だっただろう。逆に、定数の値が1/137より小さかった場合、化学反応が安定せず、化学反応を通じて生成される分子や物質が不安定で簡単に崩壊する現象が起こるだろう。このような不安定な分子では、DNAとタンパク質で構成される生命体を作ることができない。
微細構造定数はなぜこのような特定の値(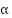 ≈ 1/137)を持ち、宇宙の秩序をよく保っているのか多くの物理学者に驚きを与えている。これについてディラックは「物理学で最も根本な未解決の問題」と言い、ファインマンは「微細構造定数は、人間が理解できないまま私たちに迫る魔法の数字で、物理学の最大の謎である。あなたは「神の手「がその数字を書いたと言うことができるが、私たちは彼がどのようにペンを押したかは知らない」と述べた。
≈ 1/137)を持ち、宇宙の秩序をよく保っているのか多くの物理学者に驚きを与えている。これについてディラックは「物理学で最も根本な未解決の問題」と言い、ファインマンは「微細構造定数は、人間が理解できないまま私たちに迫る魔法の数字で、物理学の最大の謎である。あなたは「神の手「がその数字を書いたと言うことができるが、私たちは彼がどのようにペンを押したかは知らない」と述べた。
微細構造定数式を別の言い方をすれば、電子の速度と光の速度の比率として表すことができる。つまり、光が電子より約137倍速いということだ。また、微細構造定数が関与する電磁力は。重力に比べて約1036倍も強く、強い力に比べると137倍弱いということになる。おそらく、創造主が宇宙を造る時、物理学の定数を個別に設定せず、最も重要な定数を先に設定し、その後で他の全ての物理学定数がそれに比例して自動的に決まるようにしたのかもしれない。その最も重要な定数が微細構造定数で、その値を1/137に設定した可能性も十分考えられる。
3章の「粒子物理学と創造論」で述べたように、宇宙のすべての物質(バリオン)は、17の基本粒子——クォーク、レプトン、ゲージボソン、ヒッグスボソン——から構成されています。これらの基本粒子はいずれも固有の質量、電荷、スピンを有しており、もしこれらの値が少しでも異なっていたなら、私たちが知る原子・分子・生命・宇宙の構造そのものは存在し得なかったでしょう。
例えば、アップクォークとダウンクォークの質量差が異なれば、陽子と中性子の安定性が崩れ、水素と原子が形成されなくなります。電子の質量が異なれば、原子の大きさや化学結合が不可能になり、安定した分子が作られなくなります。また、ヒッグスボソンの質量が変化すれば、すべての粒子の質量分布が変わり、宇宙の構造自体が変化します。また、陽子と電子の電荷が異なれば、電気的に中和された原子が存在できず、クォークの電荷が異なれば陽子と中性子の性質が変わり、原子核構造が不可能になります。さらに、電子がスピン1/2でなければパウリ排他原理が成立せず、原子構造を維持できません。ボソンのスピンが異なれば、電気力・強力・弱力などの力の伝達体系が崩壊します。最後に、ヒッグス・ボソンがスピン0でなければ、質量付与メカニズムが機能せず、粒子は現在のように形成されません。
このように、宇宙の始まりから質量密度、基本物理定数、そして17の基本粒子の固有の特性に至るまで、すべてが精密に調整されていることがわかります。臨界質量密度の絶妙な一致、重力・プランク・微細構造定数などの物理定数の繊細な値、そしてクォーク・電子・ボソンの質量と電荷、スピン適切な配置は、すべて生命と宇宙の存在を可能にする必須条件である。もしこれらの一つでも現在と異なっていたなら、原子や分子は安定して存在できず、星や銀河が形成されることもなく、したがって生命体も存在できなかったでしょう。これは、コンサートで全ての楽器がオーボエの「ラ(A)」の音に合わせ完璧な調和を成すように、創造主の精密な設計と意図の下、全ての物理定数と粒子が微細に調整されているため、現在の宇宙が維持され、私たちが存在できることを示しています。
このように、創造主の作品がどのように生まれ、どのような原理で機能するのか——重力、相対性理論、不確定性原理、パウリ排他原理、ヒッグス機構など——その一部を発見するだけで、世紀の天才と呼ばれたりノーベル賞を受賞したりします。では、これらすべてを最初から設計し完成させた創造主の神は、どれほど偉大なのでしょうか?
2. 神様の傑作 地球
私たちが住んでいる地球は、人間の生存に必要な水、空気、適切な気候など、生存に不可欠なすべての要素を提供している。しかし、同じ地球型惑星である金星や火星の場合、水と空気がほとんどないだけでなく、気温が極端に高かったり低かったりするため、生命体が生きることが難しい。しかし、さらに深く探求すると、地球は単に生存に不可欠な条件を備えているだけでなく、生命の存在と持続的な保存のために様々な面で極めて精密に調整された特別な惑星であることが分かる。これらの条件は非常に精巧に調整されており、まるで微調整された宇宙の延長線上にあると言える。この章では、地球が宇宙で非常に特別な惑星である10の理由を探ってみたい。
a. 太陽からの適度な距離
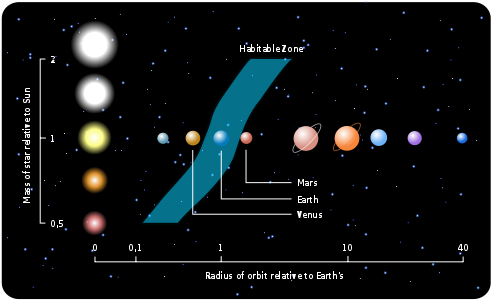
私たち人間をはじめ、地球上のすべての動植物は、水がなければ生存することができない。しかし、生命に不可欠な水も、惑星の温度が低すぎて凍った状態や、逆に温度が高すぎて沸騰した状態である場合、生命体の生存には不適当である。惑星がその中心にある星から近すぎて水が沸騰したり、または遠すぎて水が凍ったりしない、適度な距離に位置して液体状態の水を持つことができる領域を生命居住可能領域(habitable zone)と呼ぶ。太陽系内での生命居住可能領域の距離を計算してみると、その範囲は0.95天文単位から1.15天文単位の間になる。1千天文単位は地球から太陽までの距離だが、偶然にも私たちの地球はこの生命居住可能領域の中心に位置している。したがって、もし地球が現在の位置より太陽に5%だけ近ければ、すべての水が沸騰し、逆に15%ほど遠ければ、水が凍り地球は生命体が存在できない惑星となるだろう。
次の図は太陽系内の生命居住可能領域を表示したもので、地球公転軌道の周りに緑色で表示された部分が生命居住可能領域である。太陽系の一番外側で公転している海王星までの黄道面上では、この生命居住可能領域が占める面積の割合はわずか0.05%に過ぎない。しかし、この図に表示された生命居住可能領域は、単に水が沸騰せず凍らない領域だけを表示しただけで、実際に人間のような高等生命体が生きるのに適した温度範囲は約5℃から50℃の間に限る。なぜなら、寒すぎたり、暑すぎたりする場合、深刻な気象災害が発生し私たちの生存が脅かされ、最終的には生存に不可欠な農作物の栽培が不可能となるからだ。
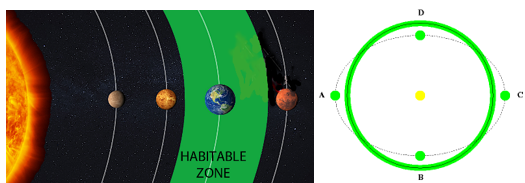
図2.1.太陽系の生命居住可能領域(緑色)
ところで、ここで重要な事実の一つは、地球が生命居住可能領域内に位置していても、地球の軌道が太陽を中心に円軌道(図の実線)ではなく楕円軌道(図の点線)で公転していた場合、地球上で生命体の生存は非常に難しかっただろうということだ。なぜなら、地球が太陽に近づくBとDの地点では地球上のすべての海水が沸騰するほどの高温になり(偏心率>0.5)、反対に太陽から遠ざかるAとCの地点ではすべてが凍るほど極寒の低温になり(偏心率>0.3)氷の惑星になるからだ。しかし、幸いなことに、地球は偏心率が0.017とほぼ円形に近い軌道で公転しているため、このような極端な気候変化がなく、生命体が安定して存在できる環境が維持されている。
b. 回転軸が適度に傾いている地球
地球の自転軸の傾きは、生物の生存に非常に重要な影響を与えている。現在、地球の自転軸が約23.5度で傾いており、この適度な傾きのおかげで、私たちは温暖な気候と豊かな四季を持つことができるようになった。しかし、もしこの地球の自転軸が直立していたり、あるいは極端に傾いていたりしたら、どうなるだろうか。その場合には、深刻な気象災害が発生し、生命体の生存を直接的に脅かすことになる。
まず、地球の自転軸が全く傾くことなく直立していた場合、どのような現象が起こるのか詳しく見てみよう。この場合、地球上のどの地点でも、一年を通じて常に同じ角度と強度の太陽の光を受けることになり、四季の変化がなくなる。例えば、次の左側の図に示されたように、赤道付近に住む人々は、地球が太陽公転軌道上のどの地点に位置していても、太陽が常に真上から照りつけるため、今よりもずっと蒸し暑い夏だけの気候を持つことになる。逆に地球の極地に住む人々は、今よりもずっと寒い冬だけの気候が続くことになるだろう。その結果、地球上で人間や他の生物が快適に暮らせる地域は中緯度地域のごく限られた範囲になるため、居住可能な空間が減り、それに伴う環境問題が深刻になるだろう。
しかし、これよりも深刻な問題は、気候変動に伴う異常気象現象である。例えば、台風は通常、海面温度が約27度以上の時に発生し、主に赤道付近の海面温度が高くなる7~8月によく形成される。台風は移動しながら海面からエネルギーを吸収して勢力を大きくするが、もし台風が通過する海域の水温が低い場合は勢力が徐々に弱まる。しかし、エルニーニョ現象などによって海面温度が高くなると、台風はさらに強力になりやすくなる。もし自転軸が直立している場合、赤道付近の海面温度は一年中高い状態が続くことになる。そうなると、台風が一年を通じて発生する可能性が高まり、しかもより強力な台風が頻繁に発生するようになる。また、海面温度が高いと、海水の蒸発が増加し、台風とともに大規模な洪水を引き起こすことになる。たとえば、韓国の場合、現在でも夏季の1~2ヶ月間に発生する台風被害によって大きな影響を受けるが、もしより強力な台風と洪水が一年中続くような気候になれば、その被害規模は想像を超えるものとなるだろう。その結果、農作物はまともに育たず、食糧不足が深刻化する。また、絶え間なく発生する洪水被害のため、人間が安定的に暮らせる地域を見つけるのも難しくなるだろう。
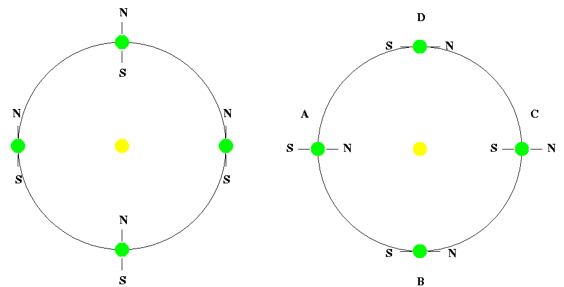
図2.2.回転軸が0度の場合と90度の場合
逆に、もし地球の自転軸が90度傾いていたらどうなるだろうか?この場合、赤道付近に住む人々は、地球が公転軌道上のA地点とC地点に位置する際、それぞれ1回ずつ、合計2回の厳しい冬を経験することになる。B地点とD地点においては、極度の猛暑の夏を2回迎えることになるだろう。また、極地方に住む人々は、6ヶ月間昼だけが続き、残りの6ヶ月間は夜だけが続く環境の中で生活しなければならない。北極地方では、地球がA地点にあるときは昼だけが続く極端に暑い夏となり、C地点にあるときは夜だけが続く厳しい冬を迎えることになる。南極地方では、北極と6ヶ月の差を置いて同じ気候パターンが現れ、A地点では夜だけが続く寒い冬を、C地点では昼だけが続く非常に暑い夏を迎えることになる。
このように昼と夜が長く続く環境では、通常の経済活動や社会活動に大きな制約が生じる。また、極地の極端な寒さによって氷河や海水が凍っていた後、6ヶ月後に猛烈な暑さの夏が訪れると、それらが急激に溶け、海水面が周期的に大きく上昇し、地球上の多くの沿岸地域が水没する。さらに、溶けた氷河の冷たい水が大量に海に流入することで、海水の温度や海流のパターンが変化し、ラニーニャ現象のような極端な異常気象が発生し、生態系の急激な環境変化をもたらす ことになるだろう。
私たちの太陽系の水星と木星の場合、自転軸がほとんど傾いていない(水星0度、木星3度)。一方、天王星の自転軸が98度傾いている。もし地球の自転軸もこのように垂直に近い、または極端に傾いていたなら、たとえ地球が生命の生存に適した生命居住可能領域内に位置していても、地球は人間が住むには過酷な環境になっていただろう。
c. 適度な回転速度および空電の速度のストリップ
人間の生体リズムとそれに伴うすべての社会活動は、私たちが住んでいる地球の自転速度に絶対的な影響を受ける。地球の自転速度は24時間で、これによって昼と夜がそれぞれ約12時間ずつ分かれている。この周期は、一日約8時間働き、8時間は余暇生活を楽しみ、残りの8時間で睡眠をとるという理想的な生活パターンを可能にする。しかし、地球とは異なり、太陽系内の他の惑星の自転速度はそれぞれ異なり、もし私たちがこのような惑星に住んでいたら、地球とは異なる生体リズムに従うことになっただろう。
例えば、木星の自転周期は地球より短い10時間程度であるため、昼と夜はそれぞれ約5時間程度になる。このような木星の自転パターンに合わせて生活する場合、約3時間程度働き、3時間余暇時間を持ち、3時間は睡眠をとることになる。退屈しなくて良さそうだが、現実的には非常に非効率的である。例えば、通勤途中の交通渋滞がひどく、会社に着く前に退勤時間が来てしまうこともあろう。家に帰って少し休もうとすると、すぐに寝る時間になり、ベッドに横になって少し寝返りを打つと、すでに朝日が昇っているというような、非常に不規則で生産性が低い惑星での生活になるだろう。さらに、もっと深刻な問題は環境的な変化だ。地球で風が形成される主な原因は、気圧の経度力、摩擦力、そして地球の自転による転向力(コリオリ力)の相互作用によるものだ。もし私たちが地球より自転速度が2倍以上速い惑星に住んでいれば、転向力は強くなり、風の強さはより強くなるだろう。特に、コリオリ力は極地より赤道地方でより強く作用するため、このような条件で台風が発生すれば、現在よりもはるかに強力な嵐が形成され、農作物の被害や洪水被害が深刻化するだろう。
逆に、もし私たちが自転速度が遅い惑星に住むことになったらどうなるのだろうか?金星は自転速度が遅く、一回の自転に243日かかる。そのため、昼と夜がそれぞれ約122日程度続くことになる。このような場合、太陽に面した側は122日間太陽にさらされ、すべてが灼熱になるくらい温度が上がるだろうし、その反対側は真っ暗な暗闇の中ですべてが凍るくらい温度が下がるだろう。こうした環境では、農作物の栽培が難しく、気象災害だけでなく、深刻な食糧難も引き起こすだろう。さらに、自転速度が遅くなることにより、別の深刻な問題が発生する。地球の内部には金属で構成された核が存在しており、地球の自転によって金属核内の対流現象が発生し、強力な磁場が形成される。しかし、自転速度があまりにも遅いと対流現象が起こらず、磁場が発生しない。この場合、太陽と宇宙から来る有害な放射線から地球を守っている磁気シールドが形成されず、有害な宇宙線に直接さらされ、生命体の生存に大きな脅威を受けることになる。
惑星の自転速度だけでなく、惑星が太陽の周りを回るのにかかる時間である公転速度も生命体の生活パターンに大きな影響を与える。地球の公転速度は365日で、これにより1年に約3ヶ月単位で春、夏、秋、冬の四季が繰り返される。
しかし、私たちが水星のように公転周期が短い惑星に住んでいたらどうだろうか?水星の公転周期は約88日で春、夏、秋、冬が22日周期で変わることになる。季節が約3週間という短い周期で変わるようになれば、急変する季節に適応するために、今よりもはるかに面倒な生活を送ることになるだろう。地球上のほとんどの農作物は、種を蒔いてから収穫までに約6ヶ月かかる。しかし、もし季節が3週間ごとに急激に変化すれば、作物が十分に熟す時間が足りないか、少し熟したとしてもすぐに冬の寒さで枯れてしまうだろう。この場合、人間の生存に直接影響する食糧不足の事態を引き起こす 。
これとは逆に、地球の公転周期が海王星のように164年になるとどうなるだろうか。この場合、春、夏、秋、冬の各季節は約40年ずつ続くことになり、人間を含む動植物の生存に深刻な影響を与えるだろう。公転周期が1年の現在の地球でも、平年より冬が長くなると、多くの動植物は厳しい寒さや食糧不足で生存に苦労するとなるが、もし冬が40倍も長くなると、その影響はさらに大きく、多くの動物が耐えられず絶滅するだろう。人間も40年間農作物を栽培することができなければ、食糧不足が深刻し生存そのものが脅かされることになるだろう。
d. 適度なサイズのストリップ
地球の大きさも、地球上に住んでいる人間の生存と密接な関係を持っている。普段、一深く考えたことのない地球の大きさが私たち人間の生存に重要な影響を与えるという事実は聞きなれないかもしれないが、地球の大きさは人間だけでなく、地球上のすべての生命体に大きな影響を与える。では、地球の大きさが生命体にどのような影響を与えるのか、一度調べてみよう。
もし地球が今より半分の大きさになったらどうなるだろうか? この場合、地球の質量は半径の3乗に比例するため、半径が半分になると質量は1/8に減り、地表面の重力は質量に比例し、半径の2乗に反比例するため、今より半分になる。私たちが毎日呼吸している地球の空気も質量を持ち、地球の重力の影響を受けることになる。 現在の地球では、地表付近には空気の密度が高く、山に上がるほど空気の密度が薄くなる。ところが、地球が半分に小さくなると、重力が弱くなるため、それに比例して地球の脱出速度も小さくなる。その結果、空気粒子の自由運動速度が地球脱出速度より大きくなる状況が生じ、大部分の空気が大気圏外に逃げてしまうことになる。こうなると、地球は空気が次第に希薄になり、空気で呼吸するすべての動植物は生存できなくなる。実際、地球の半分の大きさの火星では少ない重力によって空気の大部分が宇宙に逃げてしまい、現在残っている空気は地球の1%程度しかなく、地球の大きさの1/4しかない月は火星よりも弱い重力で、遥か昔に空気が宇宙の外に失われてしまった。
重力が少なくなると、空気が減少する以外に私たちの身体にも変化が起こる。宇宙飛行士が重力の弱い宇宙空間に行くと関節が伸びて身長が少し高くなるように、人の身長は弱い重力によって今より大きくなるだろう。また、歩行は重力の少ない月でのアポロ宇宙飛行士のように、ガンガン飛び跳ねる歩行になるだろう。想像してみよう。マッチ棒のようにガリガリに細くなった体を持った人がウサギのようにぴょこぴょこ跳ねていたら、どれほど滑稽に見えることだろうか。しかし、幸いなことに、創造主である神様は私たちを、美しく神様の形に従って創造された。そして、このような滑稽な姿にならないように、地球も私たちにとって最適な大きさに作られているのだ。
逆に、もし地球の大きさが今よりもっと大きかったらどうなっていたのだろうか?地球の大きさが大きくなることで生じる問題も、重力に関係している。地球の重力が大きくなると、水素やヘリウムのような軽い気体だけでなく、体に有害なアンモニアやメタンといった気体も宇宙に逃げることができず、大気にどんどん溜まってしまう。こうなると、重力が大きい木星のように地球の大気成分が変化し、生命活動に必要な酸素ではなく、体に有害なメタンなどの有毒ガスが蓄積され、生存そのものが不可能な状況になる。
地球の大きさが大きくなることで生じるもう一つの脅威は、強くなった重力によって今よりもはるかに多くの隕石を地球に引き寄せ、私たちの生存に直接的なダメージを与えるということだ。もし隕石が現在よりも多く落ちるようになれば、隕石がどこに落ちるかについての隕石予報が天気予報よりも大きな比重を占めるようになり、外出時には、すべての人が厚いヘルメットを着用し、家や車も隕石に備えて厚い鉄板で武装する必要があるだろう。 また、地球の重力が大きくなれば、重い重力に耐えて歩くために、すべての人が象足のように丈夫な脚を持ってウロウロ歩くことになり、身長もよく伸びにくくなるだろう。
地球の大きさの変化に伴う酸素量の変化も、人間の生存に大きな影響を与える。もし現在、大気の21%を占めている酸素の量が今より少なくなったらどうだろうか。そうなれば、当然、呼吸が困難になり、多くの酸素を必要とするスポーツ活動は決してできないという制限的な生活を送ることになるだろう。例えば、険しい野山に登る際に、酸素ボンベを背負って行かなければならないかもしれない。
逆に、もし酸素が今より多く存在したらどうなるだろうか? この場合、新鮮な酸素をたくさん吸えることは一見良いことのように思えるが、多くの酸素によって地球は常に火事に見舞われることになるだろう。日常生活においても容易に火花が発火し、どの公務員よりも消防公務員が必要になるだろう。酸素が多いことで生じるもう一つの問題は、酸素原子が3つ集まったオゾンが過剰に生成されることだ。オゾンを呼吸中過度に吸い込むと、肺に損傷を与え、特に呼吸器疾患のある人々には致命的になる。このような理由でオゾン注意報が頻繫に発令され、外出時には誰もがオゾンを浄化してくれるマスクを着用しなければならない不便な世の中になるだろう。
図2.3.太陽系惑星の大きさの比較
e.強い磁場を持っている地球
地球は宇宙空間から絶えず降り注ぐ宇宙線(Cosmic ray)にさらされている。そのほとんどは、太陽から太陽風のように放出される荷電粒子や超新星の爆発によって生じた粒子である。もしこれらの粒子がろ過されずに地球に直接入ってくると、地球上の生命体の生存に深刻な影響を及ぼすことになる。
その影響の一つ目は、荷電粒子の高いエネルギーによって細胞が破壊されたり、突然変異を起こしたりすることである。二つ目の影響は、このような荷電粒子がビリヤードボールが他のビリヤードボールを押しのけるような打撃作用(Sputtering)を引き起こし、この作用で地球の上層大気の一部がを宇宙空間に吹き飛ばされ、空気が減少することである。私たちが呼吸するために必要な空気が減少することは、どれほど深刻な問題なのだろうか。実際、火星の磁場は地球の磁場の万分の一程度しかなく、このような弱い磁場により、わずかに残っている二酸化炭素で構成された大気も、この打撃作用によってますます希薄になっている。しかし、幸いなことに、私たちの地球はこのような危険な現象を防ぐ効率的な 磁気シールド膜を持っている。
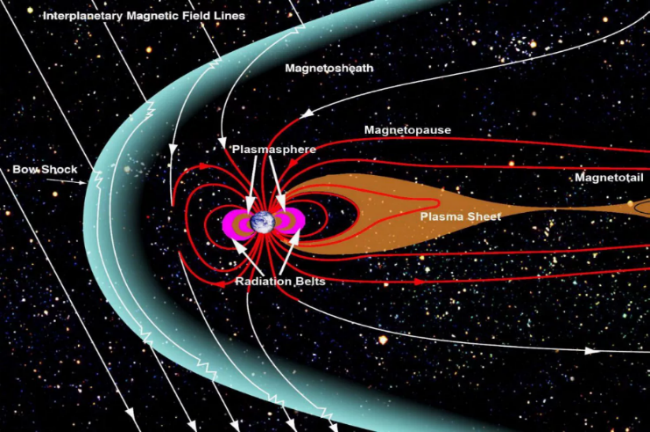
図2.4.地球を包み込む磁気シールド膜
地球の内部には鉄とニッケルで構成された液体状態の金属核があり、この金属核が地球の自転によって対流現象を起こすと、強力な磁場が発生する。こうしてできた地磁気によって次の図に示すように、地球全体を包み込む巨大な磁場が形成され、太陽活動によって太陽風と共に吹き込む荷電粒子が地球の近くに近づくと、この巨大な磁気圏によって地球上に直接入ってくることなく、磁気圏界面に沿ってずれていくようになるのだ。ただし、地球上には磁気シールドが少し弱い場所があり、代表的な場所が南極と北極地方である。荷電粒子がこれらの極地に入ると、上層大気と衝突して放電を引き起こし、その結果、色とりどりに輝くオーロラとして現れる。
このような強力な磁気シールドがなければ、高エネルギーの宇宙線がろ過されることなく地球に入ってきて細胞を破壊し、地球の空気を宇宙に逃がし、生命体の生存に致命的な影響を与えただろう。
f. 例外的に大きい月を持っている地球
月は地球上の生命体の存在において重要な影響を与えている。私たちが夜空でいつも見てきた月がどのように私たちの生存と関係があるのかを疑問に思うかもしれないが、月には主に2つの重要な役割がある。一つ目は地球の自転軸を安定させ、異常気象を防ぐことで、もう一つ目は潮汐力を発生させて海洋生態系を維持することである。
まず、月による地球の自転軸の安定化について見ていこう。私たちが先に考察したように、地球の自転軸は23.5度傾いており、地球上の生態系が快適に生きられる季節の変化をもたらしている。 しかし、地球の自転軸がこのように23.5度で一定に保たれることができる理由は、地球の近くにある例外的に大きな月の存在によるものである。 太陽系内の地球型惑星において、私たちの地球のように大きな月(半径=1,740km)を持つ惑星は存在しない。同じ地球型惑星である水星や金星には月が全く存在せず、火星にはギリシャ神話に登場するフォボスとダイモスの名を持つ2つの月があるが、その半径は月の0.6%にあたる約10kmと非常に小さい。 先に論じたように、太陽系における生命体居住可能領域は1天文単位付近にある。
一方、太陽系の惑星が通過する黄道面は海王星まで約30天文単位にわたっている。この距離を考慮すると、地球は太陽に近すぎる。 これほど近くにいなければ生存に適した温度を得られないが、一つの問題が存在する。それは地球に及ぼす太陽の重力が強すぎて、地球の自転軸を不安定にするという点だ。 太陽の重力による自転軸の揺れ現象は近隣惑星でも観測される。下図は大きな衛星を持たない火星において、太陽の重力によって生じる自転軸と離心率の変化を示している。 図を見ると、火星の自転軸と離心率は約15万年の周期で大きく振動し、過去600万年間で自転軸は15度から45度まで、離心率は0.01から0.11まで変化したことがわかる。
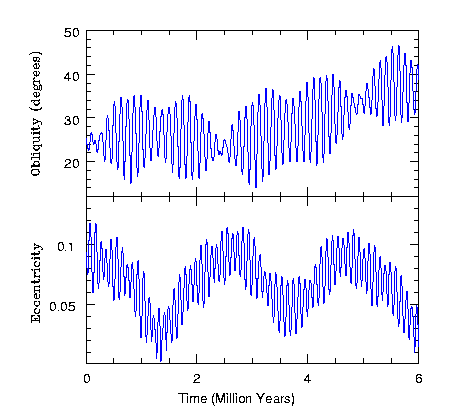
図2.5.火星の自転軸と偏心率の
地球と似た大きさの姉妹惑星である金星では、月がないため太陽の重力によって自転軸の変化が0度から180度までカオティックに変化する。 もし地球に月がないと仮定した場合、予想される自転軸の変化はどうなるだろうか?論文によって異なるが、一般に数百万年の間に約0度から70度程度の変化をもたらすと言われる。 この程度に自転軸が変化すれば深刻な気象災害を引き起こし、究極的には生命体の生存に致命的な結果をもたらす可能性がある。 しかし幸いなことに、創造主は地球の傍らに例外的に大きな月を準備しておられ、こうした災害を防がれた。
では他の地球型惑星とは異なり、地球はどうしてそれほど巨大な月を衛星として持つことができたのだろうか?月の起源についてはいくつかの仮説が提示されてきた。 火星サイズの天体が地球と衝突して月が形成されたという衝突説、別の惑星を重力で捕獲したという捕獲説、地球と月が同時に形成されたという同時形成説、そして地球の一部が分離して月になったという分裂説などである。 しかし最近の研究は衝突説を最も有力な説明として支持している。
ところが巨大衝突を通じて地球–月システムのような大型衛星が形成されるには、複数の条件が精密に満たされなければならない。
第一に、衝突体の大きさと衝突角度が非常に精巧に一致しなければならない。 衝突体(テイヤ)は、地球を完全に破壊することなく、マントル物質を宇宙空間へ十分に噴出させるために、地球質量の約10%程度の大きさでなければならず、必ず斜めの角度で衝突しなければならない。 正面衝突では月を形成する材料を生み出せない。
第二に、噴出した残骸物質は急速に凝縮・合体しなければならない。残骸が地球の強力な潮汐力によって分散されるか、再び地球に吸収される前に、短い時間的窓(accretion window)の中で月を完成させなければならない。
決定的に、月の安定した形成には衝突直後の地球の自転速度が十分に速くなければならなかった。なぜなら衝突で噴出した物質が地球に再落下せず安定した軌道を形成し、ロッシュ限界の外側で月として凝集する必要があるからだ。
このように大きさ、角度、速度、時間的条件がすべて完璧に一致しなければならないため、他の地球型惑星でこれほど巨大な衛星が形成されることは極めて稀な現象であると推定される。
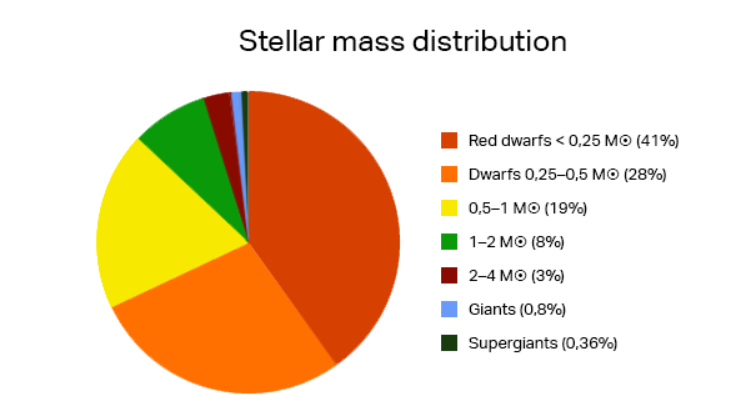
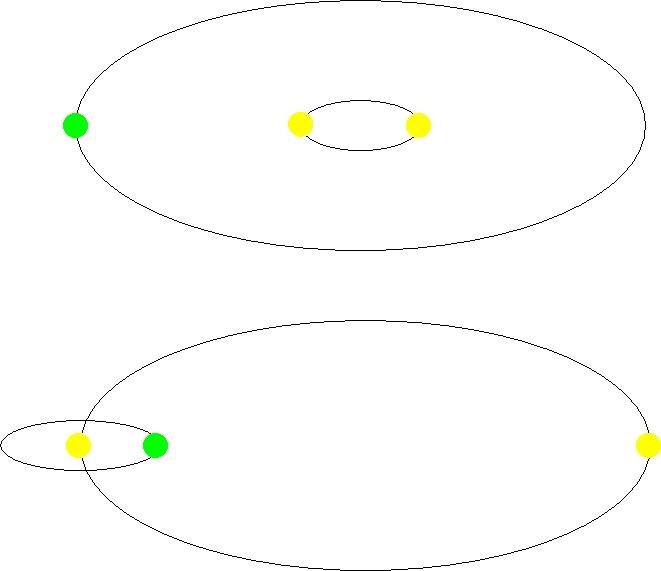
それでは、月の潮汐現象が海洋生態系にどのような影響を与えるのか見てみよう。月の潮汐現象は、満潮と干潮を発生させ、それに伴う海流の動きを引き起こす。この海流は、干潟で生活する魚介類に豊富な酸素と栄養分を伝達する。さらに、海洋生態系の食物連鎖構造の最も基本的な生物であるプランクトンに豊富な酸素を供給し、同時に栄養分が豊富な下層の栄養塩を吸い上げる竜巻のような上昇流を発生させて栄養分も提供する。また、遊泳力がない、あるいは遊泳力が微弱なプランクトンを様々な場所に運び、それを餌とする多くの魚たちに安定した食料源を供給するなど、海流は極めて重要な役割を担っている。 満潮と干潮のもう一つの重要な役割は、川から流れ込んだ栄養分の多い淡水を海水と混ぜる働きだ。もし月による潮汐の差が存在しなければ、塩分濃度の高い重い海水は下に沈み、川から流入する淡水は上に溜まり成層化現象が起こる。このような成層化が起こると、上層に集中した栄養分を利用して赤潮プランクトンが大量繁殖し、次の図のように赤潮が発生する。また、その猛毒な赤潮を食べた魚介類は斃死し、大量繁殖したプランクトンの残骸が海の底に積もると、細菌がこれらを分解し、ほとんどの酸素を枯渇させる。大量繁殖したプランクトンは魚の鰓に付着して窒息させたり、鞭毛藻類プランクトンであるココリディニウムは毒素を放出して魚を殺すこともある。 また、潮汐現象がなければ、わずか残っている上層部の酸素が下層に伝達されず、下層に住む魚の大量死をもたらすが、このような場合、沿岸部に住む小さな魚を餌にする大きな魚も食料を失い、最終的には海洋生態系の大崩壊につながる可能性がある。実際に、夏の梅雨期の多雨で河川水が大量に海に流入した後、赤潮現象が起こるのもこの成層化現象によるものである。潮汐の差が大きいところや潮の流れが速い海域では河川と海水がよく混ざり赤潮はあまり発生しないが、島が多く、水の流れが遅い海岸では赤潮現象がよく見られるのもこのような理由からだ。 図2.6.赤潮現象によって赤くなった海 このように、月は地球の自転軸を安定させ、海洋生態系を維持する重要な役割を果たしている。しかし、たとえ地球が月を持っていたとしても、その大きさが現在よりも小さかった場合、地球の自転軸を安定させる効果が十分ではなく、潮汐力も不十分だっただろう。逆に、月が今よりも大きかった場合、地球と月の間の重力が強くなり、月が地球に引き寄せられて衝突する危険性が生じる。または、大きくなった月の強い重力によって潮汐力が増加し、満潮のたびに海岸沿いの多くの都市が水没するような状況も発生しただろう。さらに、現在と同じ大きさの月であっても、今より近い距離に位置したり、もっと遠くに位置したりした場合でも、やはり同様のさまざまな問題が発生しただろう。このように私たちの生存に重要な月も、また、もし一つではなく二つ存在していたとしたら、別の脅威になっただろう。増加した潮汐力によって数十から数百メートルの高さの巨大な波が形成され、津波が発生していたかもしれない。また、それだけでなく月の強い重力の影響により、地球の火山活動や地震の頻度が増加し、地球の環境ははるかに過酷なものになっていたことだろう。地球が例外的に大きな月を、地球からちょうどよい距離に一つ持っていて、地球の自転軸が安定し、海洋生態系に不可欠な潮汐力が生まれているとは、なんとありがたいことだろうか。 g. 木星に守られている地球 木星は太陽系最大の惑星であり、地球より約11.2倍大きく、質量は約318倍重い。木星の存在が私たちの生存にどのような関連性があるのか、疑問に思うかもしれないが、木星も月と同様に私たちの生存にとって極めて重要な役割を果たしている。私たちの太陽系には火星と木星の間、そして木星の軌道付近に小惑星帯が存在するが、軌道を逸脱した小惑星と周期的または非周期的に太陽を訪れる彗星が地球に大きな脅威を与える可能性がある。このような小惑星や彗星、またはそれらの壊れた破片が地球上に落ちることを隕石と呼ぶが、今この瞬間にも多数の隕石が地球の大気圏に突入している。下の図は、地球に落ちる隕石の大きさと、その頻度を示す図である。 例えば、直径1メートル程度の隕石は1時間に1個程度、数メートルサイズの隕石は1日1個程度、そして数メートルから10メートルサイズの隕石は1年に1回程度の頻度で地球上に落下する。大気圏に突入する隕石の直径が10メートル以下の場合、大部分は空気との摩擦によって大気中で燃え尽きてしまい、実際に地表に何らかの衝撃を与えることは少ないが、問題はそれ以上の大きさの隕石が地球に落下する場合である。 図2.7.地球上に落下する隕石の大きさと頻度数 このような隕石が地表に落ちると、深刻な災害をもたらす。実際、1908年にソ連のツングースカ地域で約60メートルサイズの隕石が落下し、空中で爆発したが、その衝撃によって約2千km2面積の地域で約8千万本の木々がなぎ倒されたという(図参照)。このような規模の隕石は、およそ1世紀に1回程度落下し、その爆発力は広島型原子爆弾の約1,000個分に相当する。 図2.8.ツングースカ隕石によって倒された木々 直径100メートルサイズの隕石は約1000年に1回、1kmサイズの隕石は100万年に1回、そして白亜紀末期の6,500万年前にメキシコのユカタン半島に落ちて地球上の恐竜を絶滅させた直径約10kmサイズの隕石は、およそ1億年に1回程度で地球に衝突する。たとえば、100メートル程度の隕石が落ちた場合、直径約1km程度の隕石ピットができ、数百メガトンのTNT爆発に相当する衝撃を与える。その衝撃は、半径15km以内の地域を壊滅させ、万が一その隕石が海に落ちた場合には、数十メートルの高さの津波が発生し海岸沿いにあるほとんどの都市が水に流される。 木星が地球上の生命体にとって重要な理由は、このように地球に大きな脅威を与える隕石を地球の代わりに引き寄せて地球を守る盾の役割を果たしていることにある。計算によると、直径1km以上の周期彗星について、木星が地球より約5,000倍ほどよく捕獲するとされる。もし木星がなかったら、このような彗星は木星の代わりに地球へと落ちて大きな被害をもたらしただろう。 図2.9.壊れたシューメーカー・レヴィ彗星と木星に衝突した様子 木星の強い重力が実際に彗星を捕獲する壮観な光景が1994年に観測された。次の図は、木星の重力によって複数の破片に割れたシューメーカー-レビー彗星が、まるで掃除機に吸い込まれるよう木星表面に衝突する場面をハッブル宇宙望遠鏡が観測した写真である。写真に茶色く見える部分が彗星の破片が衝突した地点であり、衝突した彗星の中で最も大きな破片の直径が約2kmほどで、もしこの破片が木星の代わりに私たちの地球に落ちていたら、北米大陸程度の広大な土地を壊滅させるくらいの威力だろうと推測される。 しかし、ここで重要な事実は、もし木星が今よりも地球から遠く離れていたり、あるいはその質量が今より少し小さかったりした場合、これほど多くの隕石を効果的に捕獲することはできなかっただろう。適度な大きさの木星が地球から適度な距離に存在し、地球を危険な天体から守ってくれるのは、驚くべきことであり、幸いなことである。 h. プレートテクトニクスによって自動温度調節される地球 プレートテクトニクスは、地殻を構成しているプレートが地球マントルの対流現象によって動き、それによって地震や火山活動が起こり、湿谷や山脈などが形成されるという説である。では、このプレートテクトニクスが私たちの生存と何の関係があるのだろうか?結論から言えば、プレートテクトニクスは地球の温度を自動的に調節し、地球上の水が凍ったり、沸騰したりしないようにしてくれる。 プレートテクトニクスがどのように地球の自動温度調節器の役割を果たしているのか見てみよう。地球の温度は様々な要因によって決定されるが、その中で最も重要な要因は、太陽から地球に到達する太陽エネルギーの強さ、その太陽エネルギーが実際に地球上にどれだけ吸収されるか、そして地球大気中の水蒸気、雲、二酸化炭素による温室効果である。太陽から届く光は、地球上の大気をよく通過するが、地表面で反射される。この反射によって光は乱反射し、波長が長くなり、大気中の温室効果ガスによって吸収される。吸収されたエネルギーの半分は宇宙に放出され、残りの半分は地球上に戻り、地球が熱くなる。 温室効果を引き起こすガスには二酸化炭素、メタン、オゾンなどがあるが、最も大きな影響を与えるのは二酸化炭素である。大気中に二酸化炭素が多ければ温室効果が大きくなり、地球全体の温度が上がる。逆に、少なければ温室効果が弱まり、地球の温度が下がる。二酸化炭素の大部分は火山活動によって噴出される。こうして噴出された二酸化炭素は、風化作用によって生成されたケイ酸塩鉱物と反応し、炭酸カルシウム(主に石灰石)を生成しながら大気中から除去される。ところが、大気中に二酸化炭素が多くなり地球の温度が上がると、ケイ酸塩鉱物の風化作用も一緒に上昇する。このように風化作用が上昇することで、より多くのケイ酸塩鉱物が生成され、大気中の二酸化炭素と反応して石灰石を作りながら二酸化炭素量を減少させる。二酸化炭素の減少により温室効果も弱まり、地球の温度は下がる。こうして地球の温度が下がると、ケイ酸塩鉱物の生成も一緒に減少し、二酸化炭素との反応が少なくなるため、大気中の二酸化炭素の割合が上がる。これにより、温室効果も強まり、地球の温度がどんどん上昇する。地球の温度が上昇すると、再びこのサイクルが繰り返されることで、地球の温度は自動的に調整され常に適正な状態に維持することになる。つまり、二酸化炭素の循環によって地球の温度は自動調節されるということだ。下の図は、過去80万年の間にこの過程がどのように作用してきたかを示している。二酸化炭素量が増加すると地球の温度が上昇し、二酸化炭素が減少すると地球の温度が低下することを示している。 図2.11.二酸化炭素増加量と地球温度の相関関係 しかし、驚くべきことは、二酸化炭素による自動温度調節作用が、プレートテクトニクスによる地殻の移動がなければ働かないという事実である。次の図のように、地球は火山活動を通じて二酸化炭素を大気中に放出し、こうして放出された二酸化炭素は風化作用によって生成されたケイ酸塩鉱物と反応して石灰石に変わり、海中の沈殿物として堆積する。海中に堆積した石灰石は、海底地殻の一枚のプレートが他のプレートの下に沈み込むサブダクション(subduction)作用によって地球内部に入り、熱い熱で溶ける過程で再びマグマの噴出とともに二酸化炭素を大気中に放出する。 もしプレートテクトニクスによるサブダクション作用がなければ、海中に堆積された石灰石は海底に堆積し続け、海中だけに留まることになり、二酸化炭素は大気中に放出されない。その結果、温室効果が減り、地球の温度は急激に下がることになる。もし歴史上、一度でも二酸化炭素循環サイクルが壊れて作動しなかったら、地球上に生命体が存在することは難しかっただろう。なぜなら、温度が下がって海水が凍る程度になると、太陽から届く光はほとんど地球に吸収されず、氷に反射されるようになる。そうなると、もう取り返しのつかない「暴走氷室効果(runaway icehouse effect)」が起こり、地球は完全に氷に覆われた惑星に変わってしまうからだ。 図2.12.摂食作用による二酸化炭素循環過程 地球の自動温度調節において重要な役割を果たすこのプレートテクトニクスがどのように始まったのか、また、どのように長い間維持されてきたのかについての研究はごく最近になってから行われている。研究が進むにつれ、プレートテクトニクスに関する神秘的な事実が明らかになり、最新の研究結果によると、もし地球の大きさが現在より20%程度大きかったり、地殻に含まれる鉄やニッケルなどの金属成分が今よりも少し多かったり、または地殻の厚さが今より厚かった場合、、プレートテクトニクスは正常に作動しなかったという。 地球が適度な厚さの地殻を持ち、その地殻がプレートテクトニクスによって徐々に移動し、わずか0.03%の二酸化炭素を大気中に循環させ、地球の温度を自動的に調節させることはいかに神秘的か。 i. 適度な大きさの太陽を持っている地球 私たちの太陽系において、水が凍ることも沸騰することもない領域を生命居住可能領域としたが、この生命居住可能領域は私たちがどの星の周りに住んでいるかによってその領域が変わる。次の図に示されたように、太陽より小さい星の周りに住むと、星の輻射熱が少なく、そのため生命居住可能領域は星の近くに位置する。反対に、太陽より大きい星の周りに住むと、多くの輻射熱のために生命居住可能領域はその星からより遠くなる。 図2.13.星の大きさによる生命居住可能領域の変化 星の質量が少なく、生命居住可能領域がその星の近くに位置する場合、潮汐ロック現象が起こる。惑星とその星が同周期自転(Synchronous rotation)をするようになる現象で、両手を合わせて向き合いながら回るようになると、片側は常に日光を受けるため、水が沸騰するほどの温度になり、その反対側は常に夜であり、凍った凍土の地となる。このような環境では、生命体の生存はほとんど不可能である。また、惑星が自転しないため、磁場を形成することができない。磁場がなければ、星から放射される有害な放射線が細胞やDNAを損傷し、生存を脅かす。 逆に、星の質量が大きくて生命居住可能領域が星から遠く離れている場合、生命居住可能領域の寿命が長く続かないという欠点がある。なぜなら、質量が大きい星は質量が小さい星に比べて核燃料である水素をより早く消費するようになり、ヘリウム燃焼を始めながら赤色巨星に進化する。こうなると星の大きさがどんどん大きくなり、近くにある惑星自体を飲み込むほどになってしまうからだ。赤色巨星に進化する星の寿命は通常数百万年に過ぎず、DNAと細胞に有害な高レベルの紫外線とX線を放出し、生命体が住みにくい環境を作る。また、星のエネルギー放出にも多くの変動があり、惑星の気候を不安定にし、極端な温度変化を起こして生命体の生存を困難にすることもある。 したがって、長い間、安全な生命居住可能領域にとどまるためには、太陽と似た大きさの星を地球のエネルギー源として持つことが必要である。その場合、赤色矮星や赤色巨星とは異なり、長い期間安定したエネルギーを提供し続けることができる。星がどのような質量分布を示すかを観測したデータを見ると、星の約88%は太陽より軽い星であり、太陽の質量の1-2倍の間にある星は8%を占め、残りの4%は太陽よりも2倍以上重い星である。したがって、太陽と同じ質量を持つ星を見つける確率はわずか数パーセント以内にすぎず、その確率は決して高くないことが分かる。 図2.14.星の質量の分布 私たちが長期間にわたって安定した生命居住可能領域にとどまるためには、太陽と似た質量の星をエネルギー源として持つことが必要であるという事実に加えて、もう一つの条件が追加される。それは、地球が連星系ではなく単星系に存在しなければならないということである。私たち人間にも双子が時々生まれるが、太陽質量の星では約40%は連星系に属しており、連星系はさらに近接連星系(Close binary system)と遠隔連星系(Wide binary system)に分けることができる。近接連星系では、惑星が回っている場合、次の図の上部のように二つの星の周りを回っており、遠隔連星系では、図の下部のように片方の星の周りを回ったり、それぞれの星の周りを回ったりする。もし私たちが住んでいる地球に太陽が二つある場合を仮定してみると、私たちの生命にとって有害な影響が多くなるだろう。まず、二つの太陽から私たちの地球に有害な放射線がより多く降り注ぐことになるだろうし、さらに、二つの太陽の相対的な位置によって地球の自転軸と偏心率が常に変動し続けて、深刻な気候及び環境変化をもたらし、生命体が住むのに適した惑星にはならない。 図2.15.近接連星系(上)と遠隔連星系(下) j. 銀河中心からの適度な距離 に位置する地球 先ほど見たように、星の周りの特別な領域でのみ生命居住可能領域が存在するように、私たちの銀河内でも特別な領域でのみ生命居住可能領域が存在する。この領域を銀河生命居住可能領域(Galactic habitable zone)と呼ぶ。 太陽系内での生命居住可能領域は、生命に不可欠な水が液体状態で存在する領域として定義されるが、銀河系内での生命居住可能領域は、生命体と地球を構成している重元素含有量が適度であるか、そして超新星やガンマ線爆発の危険からどれだけ安全な領域に位置しているかというのと関連している。天文学で定義される重元素とは、水素やヘリウムより重い元素を指すが、これらの重元素の含有量は銀河内でその分布が均一ではなく、銀河中心からの距離によって異なる分布をみせる。 生命体が維持されるためには、その生命体が摂取する食品にその生命体の構成成分に必要な栄養素が含まれていなければならない。また、これらの食品は土からその食品自体が成長するために必要な栄養素を取ることが求められる。したがって、私たちの地球を構成する物質は、地球上の生命体が必要とする有機物を必ず含んでいなければならないが、この有機物を構成している構成成分のほとんどは炭素、窒素、酸素などの重元素である。さて、もし私たちの太陽系が現在の位置より銀河の中心から遠く離れていたとしたら、重元素の含有量が不足し、生命体が維持できなかっただろうし、逆に、銀河の中心により近づいていたら、あまりにも多くの重元素含有量によって生命体の存在に逆効果をもたらしただけでなく、地球のプレートテクトニクスがうまく機能しなかっただろう。このような理由から、銀河系内での太陽系の位置は、私たちの生命体が存在し続けるため重要な関連性を持っている。 私たちの地球に適量の重元素が存在するもう一つの理由は、地球内部に存在する放射性同位元素の含有量が関係している。地球は太陽から受け取る熱エネルギーだけでなく、地球内部に存在するトリウムやウランなどの放射性同位元素の崩壊で発生する熱エネルギーによっても温められ、温度を維持している。もし地球内部にこのような放射性同位元素がなかったら、地球はとっくに冷えてしまい、生命体が存在する惑星にはならなかっただろう。逆に、地球内部にあまりにも多くの放射性同位元素が存在していたら、強い放射線はもちろん、さらに過度の熱エネルギー生じて海水が沸騰し、生命体が生きられない環境になっただろう。このような重要な役割をする放射性同位元素の分布も銀河内の重元素含有量と同じようなパターンを示す。もし地球の位置が銀河の中心から外角に向かってより遠く離れていたなら、地殻内の放射性同位元素の量が少なく、地球が冷えてしまい、逆に銀河の中心に向かって近すぎたら、放射性同位元素の量が過剰になり地球の温度が高くなりすぎて、どちらこそ生命体が住めない惑星になっただろう。 私たちの地球が今より銀河の中心に近く位置したときに生じるもう一つの問題は、銀河の中心に集まっている多くの星のために発生する。地球が属する太陽の周りに星が多い場合、その周囲の星々による重力摂動で地球の軌道や自転軸が揺れ、最悪の場合、地球が太陽系外に飛び出してしまう危険性もある。近くの星の重力摂動は、オルト雲とカイパーベルトにある天体の軌道を乱し、より多くの数の彗星が太陽系に流入し、地球と衝突する可能性を高める。また、銀河の中心は星の密度が高く、近距離で発生する超新星爆発やガンマ線爆発によるガンマ線やX線も地球上の生命体に深刻な脅威を与える。もし私たちの太陽系から約1万光年以内でガンマ線爆発が起これば、相対論的な速度の高エネルギー粒子が地球上に降り注ぎ、私たちの地球を包んでいるオゾン層をイオン化させて酸化窒素化合物に転換させ、約40%のオゾン層を破壊する。オゾン層が破壊される場合、太陽から来る紫外線(UVB)がろ過されずに大気を通して入ってきて、私たち人間を含むすべての生命体のDNAを破壊して突然変異や皮膚癌を引き起こす。40%程度のオゾン層が破壊されると、DNAの損傷率が現在より約16倍まで増加し、海洋生態系の食物連鎖の基本を形成している植物性プランクトンをすべて全滅させることができる量以上の紫外線が降り注ぐ。 図2.16.植物性プランクトン(Phytoplankton) この植物プランクトンがなくなれば、海洋生態系は崩壊し、それに伴い陸上の生物も時間が経つにつれて有害な紫外線の影響でどんどん個体数が減少していくだろう。最近の研究結果によると、この植物性プランクトンが地球温暖化の主犯である二酸化炭素の循環において非常に重要な役割を果たしていることが明らかになった。植物プランクトンの光合成作用によって除去される二酸化炭素の量は、陸上にあるすべての木や草が除去する二酸化炭素の量と同じくらいだという。こうして除去された二酸化炭素は海の深い海底に保存されるが、もし強い紫外線で植物性プランクトンが絶滅すれば、大気中の二酸化炭素量が急激に増え、深刻な温室効果を引き起こし、地球の温度が急激に上昇するだろう。このように深刻な影響を及ぼすガンマ線爆発が実際に地球の歴史上で起きたことがあり、今から約4億5千万年前のオルドビス期末には、海洋生態系の約60%が絶滅した。この絶滅は、地球から約6,000光年離れた場所で起きたガンマ線爆発が原因とする研究結果が発表された。 前述したように、重元素含有量が多すぎず少なすぎず、超新星やガンマ線爆発の脅威が少ない銀河生命居住可能領域の領域を計算した論文がサイエンス誌に発表された。その研究によれば、その領域は銀河中心から約2万3千光年から2万9千光年の範囲に位置すると推定された。偶然にも、私たちの地球がある太陽系は銀河の中心から2万6千光年離れており、この距離は銀河の生命居住可能領域の正中央になる位置でもある。さらに、太陽系の回転速度と銀河の回転速度が一致する同回転半径(Co-rotation radius)を考慮すると、銀河生命居住可能領域は次の図で示される緑色の領域に該当する。なぜなら、この半径内にある星々だけが銀河の回転によって発生する密度波に流されることなく、安定した環境を維持できるからである。 図2.17.銀河サバイバル隊 この章では、地球だけが持つ10の特別な条件を通じて、人間をはじめとする様々な動植物が生存できる絶妙な環境について見てきた。これらの条件が一つでもずれていたら、地球は人間が正常に生活を営むことができない極限環境となり、あるいは生命体が生きられない惑星として残っていたかもしれない。そして、これらの条件が非常に精巧に調整されていることから、偶然に生成された可能性は極めて稀であることが分かる。これらの事実は、地球が単純な偶然の産物ではなく、生命体の生存と保存のために創造主によって設計された非常に特別な惑星であることを示唆する。次の章では、私たちの存在自体が単純な偶然の結果なのか、それとも神の創造の摂理によるものなのか、詳しく調べてみよう。
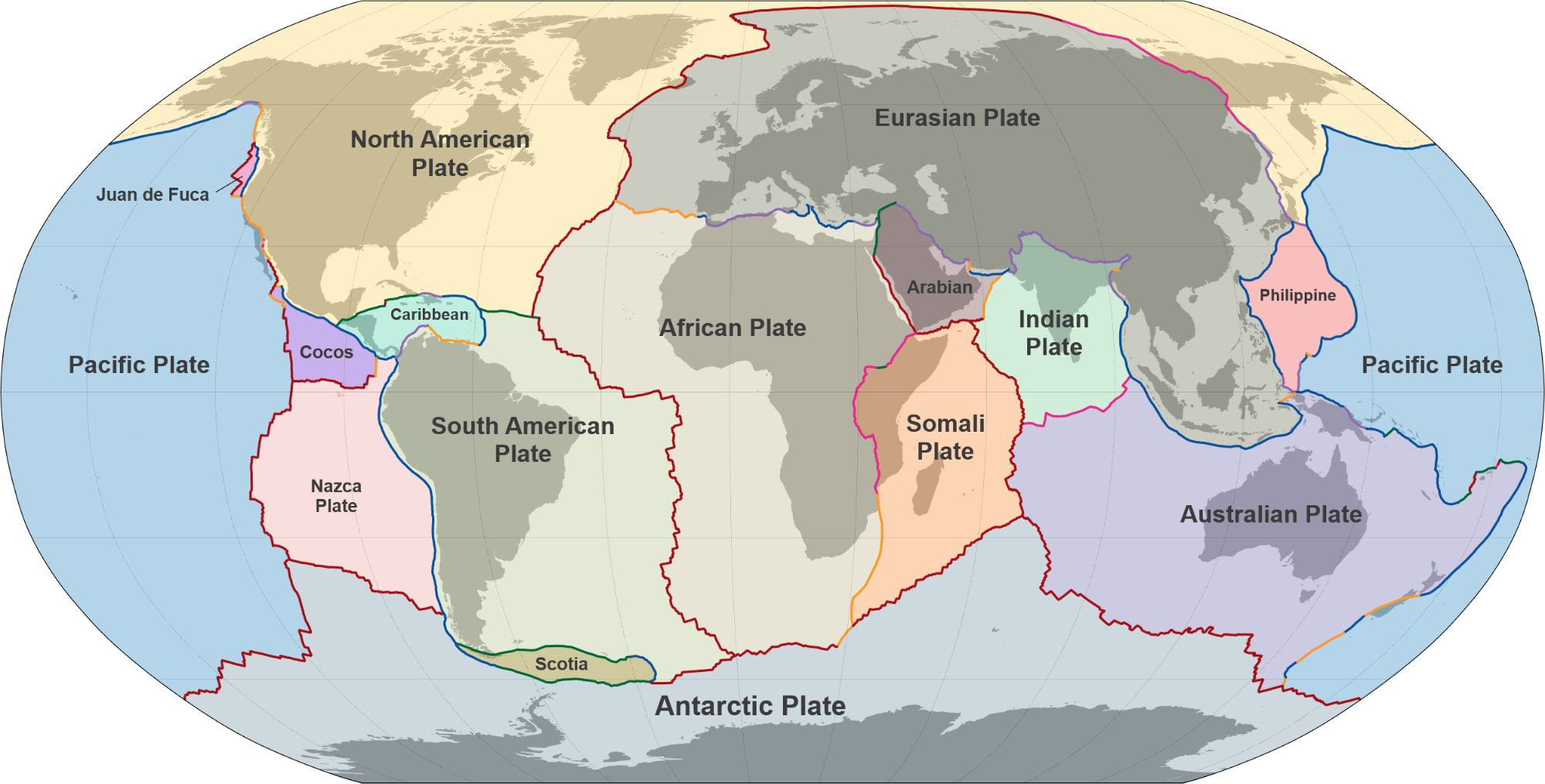
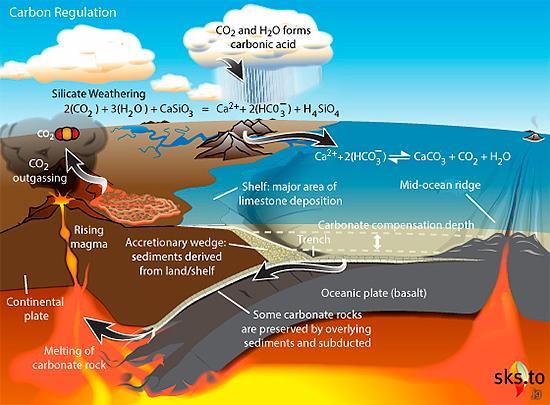
図2.10.地殻を構成しているいくつかのプレート

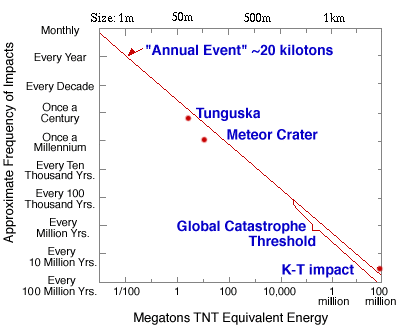


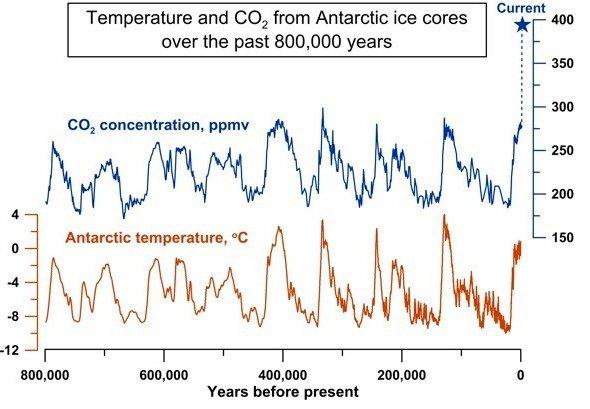

3. 創造か、進化か?
生命体はどのようにして地球上に現れたのか?それは単なる偶然による進化の結果なのか、それとも創造によるものの結果なのか?この章では、進化論と創造論に関する様々なトピックを取り上げながら、 進化が正しいのか、それとも創造が正しいのかを考えてみたい。まず、生命体が地球上にどのようにして現れたのかという生命の起源を詳しく見ていく。そして、ダーウィンの理論を「進化論」と呼ぶべきか、それとも「遺伝的適応論」と呼ぶべきかを問いただす。さらに、人間は本当に類人猿から進化したのか、知的設計論とは何か、粒子物理学、地球外生命体の存在の可能性、動物の本能、そして自然界に現れる数学的原理などを通じて、進化論の妥当性を検証していく。
a. 最初の生命体は地球上でどのように出現したのか?
最初の生命体が地球上にどのように現れたかによって、創造論と進化論の真偽が決定される。生命体が自然に現れたのであれば進化論が正しいのであり、創造主の介入なしには決して生じ得ないのであれば創造論が正しいであろう。
すべての生命体は細胞で構成されており、細胞は核酸(DNA・RNA)、タンパク質、脂質、炭水化物などで構成されている。タンパク質はアミノ酸がペプチド結合で重合されて作られ、アミノ酸はDNAに記録された遺伝情報に基づいて作られる。脂質と炭水化物もまた、DNAの遺伝情報によって作られた酵素によって生成され、これらの酵素もまたタンパク質で構成されている。したがって、生命体の基本単位である細胞が形成されるためにはタンパク質が不可欠であり、タンパク質が形成されるためにはDNAとRNAが既に存在していなければならない。しかし問題は、初期の地球にDNA、RNA、タンパク質、酵素などが全く存在しなかったという点である。つまり、生命体の始まりそのものが自然発生することは全く不可能だということである。
それにもかかわらず、進化論者たちは初期地球においてRNAがまず自ら形成され、メッセンジャーRNA(mRNA)として遺伝物質の役割を果たし、リボソームRNA(rRNA)としてタンパク質合成に関与するリボソーム酵素の役割も果たし、タンパク質とDNAを作り出したという「RNAワールド仮説」を主張する。
しかし実際には、RNA分子自体が極めて複雑で精巧な配列を必要とするため、無作為に自然生成される確率は事実上ゼロに近く、仮に偶然生成されたとしても化学的に非常に不安定で瞬時に分解される。さらに本質的に、RNAが担える触媒機能は真のタンパク質酵素に比べ極めて限定的で効率が低い。さらに、タンパク質がなければRNAは作られず、RNAがなければタンパク質は作られないため、RNAワールド仮説は構造的矛盾を抱えている。
さらに、仮にRNAとタンパク質、DNAが偶然すべて生成されたとしても、これらが自ら組み立てられて生きた原核細胞を形成し、続いて真核細胞へ進化し、細胞小器官が局在化し、細胞分化が起こり、組織・器官が形成され、最終的に多細胞生物が出現するすべての段階は、それぞれ天文学的な確率の連続である。
したがって本章では、生命体の自然発生可能性を論じるために、まず生命起源研究で頻繁に言及されるミラー・ウレイ実験について考察した後、RNAの形成、タンパク質の形成、DNAの形成、原核細胞の形成、真核細胞の形成、細胞小器官の局在化、細胞分化、組織と器官の形成、多細胞生命体の形成といった主題を探求していく。
i. ミラー・ウレイ実験
ミラー・ウレイ実験は、原始地球環境においてアミノ酸などの生命構成有機物が無機物から自然に合成される可能性を確認するために実施された実験である。
この実験では、彼らは地球の原始大気がメタン、アンモニア、水素、水蒸気で構成されていたと仮定した後、この混合ガスを加熱装置、真空ポンプ、放電装置、冷却装置などで構成された実験装置に入れて反応させた結果、四種類のアミノ酸を得た。
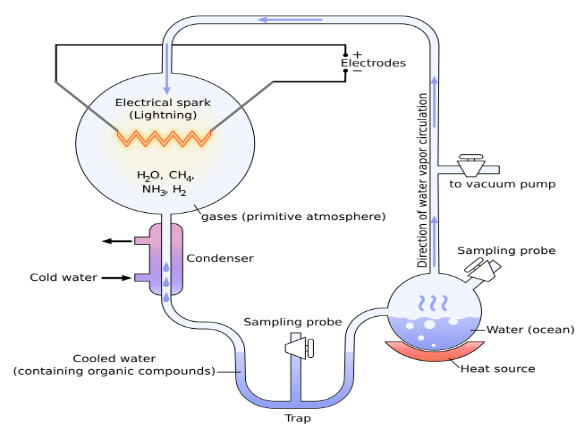
図3.1.ミラー・ユーリー実験装置
この実験に使用された加熱装置は水蒸気を発生させ、真空ポンプは空気を取り除き、放電装置は雷を模倣し、冷却装置は生成物を急冷させる装置である。 ミラー・ユーリーの実験は、ほとんどすべての生物学の本で批判なく進化論の基礎として受け入れられているが、この実験がすべての人に認められるためには、重要な検証を経なければならないと思う。 つまり、彼らの実験が地球の原始大気を正確に再現しているかという検証である。 そのために、彼らが使用した実験装置と実際の地球の原始大気を比較してみよう。
ミラー・ユーリーが自然界にある雷を模倣するために使用した放電装置は、約5万ボルトの電圧を使用し、約250度の熱を発生させる。それに比べて、雷は約1億ボルト程度の電圧を持ち、約5万程度の熱を発生する。 この比較から、ミラー・ユーリーの放電装置の温度は、雷によって発生する温度と著しい差(約200倍)があることがわかる。また、彼らの放電装置は放電が周期的に発生し、実験者は混合ガスで化学反応が起こるように放電の頻度と持続時間を人為的に制御することができた。 一方、雷は不規則であり、その持続時間は数マイクロ秒からミリ秒と比較的短い。この比較から、ミラー・ユーリーの放電装置と自然界の雷は基本的に非常に異なることが分かる。
地球の原始大気を模倣するために彼らが使用した混合ガスはどうだろうか?地球の原始大気の構成成分は、彗星の構成成分を分析すればわかる。なぜなら、彗星は太陽系を作り、残った物質だからだ。下の表に示すように、彗星の構成要素を分析した論文を調べてみると、最も多くの部分を占めるのは水(86%)であり、次に二酸化炭素(10%)、一酸化炭素(2.6%)であり、アンモニアとメタンはそれぞれ1%未満と非常に少ない。ところが、ミラー・ユーリーが使用した混合ガスは、彗星で2、3番目に多い二酸化炭素と一酸化炭素が抜けている。この結果は、彼らが使用した混合ガスが原始大気の構成成分と異なることを示している。
構成 | 比率(%) | 参考文献 |
水(H2O) | 100(86%) | Pinto et al.(2022) |
二酸化炭素(CO2) | 12(10%) | Pinto et al.(2022) |
一酸化炭素(CO) | 3(2.6%) | Pinto et al.(2022) |
アンモニア(NH3) | 0.8(0.7%) | Russo et al. (2016) |
メタン(CH4) | 0.7(0.6%) | Mumma et al. (1996) |
表3.1.彗星の組成(水=100)
ミラー・ユーリー実験のもう一つの問題は、地球の原始の大気が還元性であったという仮定である。ここで還元性大気とは、酸素が大気中に存在しないことを意味している。もし原始酸化性であったなら、アミノ酸は生成されるとすぐに酸化されて分解されてしまう。原始還元性か酸化性かによって、生命体の最も基本的な物質であるアミノ酸の生成が左右されるため、原始大気について多くの研究が行われてきた。
これまで知られていることを見ると、Miller(1953)とChyba & Sagan(1997)は還元性雰囲気、Albeson(1966)、Pinto et al. (1980)、Zahnle(1986)等は弱還元性雰囲気、そしてTrail et al. (2011)は酸化性雰囲気であると主張している。ここで注目すべき論文は、2011年にNatureに発表されたTrail et al.の論文であるが、彼らは明王帯(Hadean)に形成されたジルコンの酸化量が、現在地球上で発見されているマグマの酸化量と同じであることを見付けた。また、ジルコンの酸化量から推測すると、地球の原始大気が初めに還元性であったとしても、地球誕生約2億年後からはもはや還元性大気ではなかったと発表した。これまでに発見された最初の原始細胞化石の時期は、地球誕生後約5億年なので、生命体が発生した当時はすでにかなりの量の酸素が地球大気に存在していたことを示している。また、水を除いた原始大気中で最も高い割合を占める二酸化炭素は酸化性ガスである。この事実は、当時の大気が酸化性であることを示唆し、アミノ酸が自然に生成される可能性が低かったことを意味する。
ミラー・ユーリー実験のもう一つの重要な問題は、アミノ酸のキラル性(chirality)である。実験室で生成されたアミノ酸は、L型とD型が約50%ずつ存在する。したがって、アミノ酸が自然に生成された場合、L型とD型のアミノ酸が約50%ずつ構成されているはずだ。しかし、地球上の生命体を構成するアミノ酸はほぼ全て(99.7%)L型アミノ酸で構成されており、D型アミノ酸は約0.3%しかない。この事実も、アミノ酸がランダムな過程によって生成されなかったことを示唆している。
現代化学と生命工学では、完全に制御された実験室環境において、高純度の出発物質、精巧な触媒、最適化された温度・圧力・pH条件を活用すれば、生命体を構成する20種のL型アミノ酸を人工的に合成することができる。しかし、こうした過程は高度に設計された条件下で特定の目的のために実施される「人為的合成」に過ぎず、原始地球の過酷で非選択的な環境下で無作為に起こる自然発生過程とは本質的に異なる。
さらに、これまで原始地球の条件を再現したいかなる実験においても、20種類のL型アミノ酸が同時に生成された事例は一度も報告されていない。さらに、仮にすべてのアミノ酸が偶然生成されたとしても、これらが正確な配列で重合して機能的なタンパク質を形成したり、あるいはRNAやDNAのような核酸を自ら作り出すことは、確率的にほぼ不可能である。
それではまず、生命起源の核心的な鍵として提示されるRNAが、果たして原始地球環境において自然生成される可能性が実現可能な水準なのか、それとも事実上不可能に近いのかを詳しく見ていこう。
ii. RNAの形成
RNAの自然形成がいかに困難かを具体的に理解するには、RNAがどのような構成要素から成るかを把握する必要がある。
RNAは数十~数千個のヌクレオチドがリン酸ジエステル結合で連結された高分子である。一つのヌクレオチドは次の三つの部分から構成されている:窒素塩基(アデニンA、グアニンG、シトシンC、ウラシルU)、五炭糖であるリボース、そして1~3個のリン酸基(phosphate group)である。
したがって、RNA分子が自然発生するためには、この三つの構成要素(窒素塩基+リボース+リン酸基)がまず原始地球環境で自然に生成・精製された後、脱水縮合反応を通じて正確な順序で数百~数千回結合されなければならない。
では、果たしてこの三つの構成要素が原始地球で無作為に生成される可能性はあるのか、そしてその確率はどの程度なのか、一つずつ検討していこう。
- 窒素塩基形成
窒素塩基は、環状構造を持つ複雑な分子である。環状構造を形成するためには、特定の化学反応、特定の反応条件、そして反応触媒が必要なため、単純な原始化合物から自然に生成されることは難しい。特定の化学反応は、炭素骨格に アミン基(NH2)が付加されるアミン化反応、酸素を除去する脱酸素反応、環状構造形成反応、窒素塩基付加反応などがある。反応条件と触媒の場合、アミン化反応では アンモニアのような窒素化合物とアルデヒドまたはケトンが必要であり、反応を促進する触媒や高温の環境が必要である。脱酸素反応では還元剤が必要で、 水素やメタンなどの還元性ガスが必要である。環形成反応では、多段階の環形成が必要で、金属イオンなどの特定の触媒、高温高圧の条件、最適なpHが必要である。窒素塩基付加反応は、様々な前駆体化合物とともに高エネルギー条件が必要である。 原始地球の大気中でこれらの反応が自然に起こるためには、適切な温度、圧力、pH、前駆体化合物の十分な濃度、触媒の存在など、様々な条件が正確に合致しなければならない。しかし、変化し続ける原始地球大気において、これらの条件が偶然に揃う可能性は極めて低い。実際、実験室で人工的に最適な環境を作っても、窒素塩基の生成収率は非常に低い。さらに、仮に自然状態で窒素塩基が形成されたとしても、UV、放射線、加水分解、酸化反応などの影響で容易に分解され、蓄積されにくいだろう。
- リボース糖の形成
五炭糖であるリボースは、ホルモース反応から始まり、ホルムアルデヒド、グリコールアルデヒド、グリセルアルデヒド、エリトロ ースを生成した後、最終的にリボースが生成される。ホルモース反応は十分な濃度のホルムアルデヒドを必要とするが、原始大気条件ではホルムアルデヒドは容易に重合したり、他の化合物と反応したりする可能性があり、安定して生成されることができない。また、ホルモース反応によって生成されるリボースは収率が低く、ホルモース反応の促進のために水酸化カルシウムのような塩基性触媒が必要であるが、この触媒が存在しなかったかもしれないし、存在したとしても、原始大気状態でこの触媒が安定せず分解される可能性がある。また、効率的なホルモース反応のためにはpHが7以上のアルカリ性条件が必要であり、適切に高い30-100℃の温度が必要である。
このような困難を乗り越えてリボースが生成されたとしても、RNA合成に有用であるためには、選択的に合成され、安定化されなければならない。しかし、ホルモース反応はリボースの選択的な生成を好まず、その結果、複数の複雑な糖の混合物を生成し、リボースのみを選択してRNA合成に利用することを困難にする。複雑な混合物からリボースを選択するメカニズムとして、ホウ酸塩鉱物などの潜在的な安定化剤が提案されているが、原始的な大気条件下でのその可用性と有効性は検証されていない。もう一つの深刻な問題はリボースの安定性である。リボースは五炭糖である糖成分であるため、化学的に不安定であり、生成されても水溶液中で容易に加水分解されたり、マイヤール反応やキャラメル化などの過程を経て分解されたりしてしまう。
- リン酸塩基の生成
原始時代にリン酸基がどのように生成されたかはよく知られていない。なぜなら、リン酸自体の生成が容易ではないからである。リン酸は一般的に燐灰石(apatite)のような鉱物で発見されるが、水によく溶けず、原始大気のような水性環境で使用可能なリン酸が比較的希少だった。また、化学反応性が低く、リン酸基の生成に大きな障壁を提供する。ヌクレオチド合成に不可欠なリン酸エステルの生成には相当なエネルギーと触媒過程が必要だが、原始地球ではこのような条件が整いにくかったと推定される。リン酸基の鎖であるポリリン酸の生成は、高い温度と特別な触媒を必要とし、生成されたとしても加水分解によって単純なリン酸化合物に分解されやすい。いくつかの実験室条件で人工的にリン酸分子の生成が可能であることを示したが、初期の地球環境を現実的に反映していない高度に制御された条件で行われた実験であった。しかし、そのような制御された実験室条件下でもリン酸分子の生成収率は低いため、原始大気 状態で生命の起源を誘導するための十分な量のリン酸生成は難しいことが分かる。
- 機能性RNAが生成される確率
これまで見てきたように、RNAヌクレオチドを構成する3つのメンバーが原始地球大気中で自然に発生する確率は低い。しかし、すべての難関を乗り越え、窒素塩基、リボース、リン酸基の生成に成功したとしても、RNA本来の機能をする機能性RNAの生成という別の重要な問題に直面することになる。RNAにはいくつかの種類がある。タンパク質合成に関与するRNA(mRNA、rRNA、tRNAなど)、転写後修飾に関与するRNA(snRNA、snoRNAなど)、調節/制御RNA(aRNA、miRNAなど)、そして寄生RNAがある。RNA分子のヌクレオチド数はその種類によって異なるが、いくつかの例を挙げると以下の通りである:
- mRNA および rRNA - 数百から数千個まで。
- tRNA - 70から90個
- snRNA - 100から300個
- miRNA - 20から25個。
複数の役割を遂行するRNA分子が平均100個のヌクレオチドで構成されていると仮定すると、RNA配列の各位置はアデニン、ウラシル、シトシン、グアニンのいずれかで満たされ、長さ100のヌクレオチド配列の可能な総数は4100であり、そのうち機能性RNAが生成される確率は1/4100(=1.6x1060)=6.2x10-61となる。この確率は、全宇宙的な時間が流れても決して起こり得ない確率であり、このように極めて低い確率は、窒素塩基、リボース、そしてリン酸基が既に存在していたとしても、機能性RNAが自発的に生成されることができないことを示唆する。
iii. タンパク質の形成
タンパク質は、アミノ酸の重合によってペプチドが形成され、特定の3次元の形で適切に折り畳まれなければ、本来の機能を持つタンパク質にならない。
アミノ酸からペプチドが形成される過程は、二つのアミノ酸が反応し、一方のアミノ酸のカルボキシル基と他方のアミノ酸のアミノ基がペプチド結合を形成することから始まる。この過程で生成される水分子は縮合反応を通じて放出される。こうして形成された分子はペプチド結合を介して連結され、この過程が繰り返されて長いペプチド鎖が形成される。この鎖をタンパク質の一次構造と呼ぶ。
一次構造のポリペプチド鎖は、アルファ螺旋とベータシートと呼ばれる二次構造に折り畳まれる。これらの構造は、ポリペプチド鎖の主鎖原子間の水素結合によって安定化される。螺旋とシートをつなぐ追加の二次構造、ターン・ループは、タンパク質の全体的な折り畳みに寄与する。二次構造はさらに、疎水性相互作用、水素結合、イオン結合、および二硫化物結合によって、特定の三次元形態(三次構造)に折り畳まれる。疎水性相互作用では、非極性鎖が水性環境から離れてクラスタリング化され、ポリペプチドが圧縮された球形に折り畳まれる。水素結合は極性鎖とバックボーンの間に形成され、折り畳まれた構造を安定化させる。イオン結合は反対の電荷を持つ鎖間の静電的相互作用でタンパク質の安定性に寄与し、ジスルフィド結合はシステイン残基間の共有結合でさらなる安定性を提供する。
複数のポリペプチド鎖(小単位体)で構成される一部のタンパク質の場合、これらの折り畳まれた単位が集まって4次構造を形成する。エラーを防止するために、シャペロンはタンパク質が折り畳まれる過程を助け、誤った折り畳みや凝集を防ぎ、正しい形をとるのを助ける。タンパク質は、最も安定的で機能的な形態を得るために、わずかに形態変化をしたり、リン酸化、糖化、切断などの化学的修飾を行ったりすることもある。
アミノ酸からタンパク質が作られる様々な過程は、非常に 複雑だが、精巧によく練られたアルゴリズムに従って行われることが分かる。この事実と、タンパク質が作られる基本設計図がすでにDNAに含まれているという事実も、タンパク質の形成が方向性のないランダムな過程ではなく、精巧な知的設計の産物であることを示唆している。
確率的に見ても、ランダムな過程でタンパク質が形成されることは難しい。一つのタンパク質に含まれるアミノ酸の数は数十から数千個になるが、小型タンパク質であるインスリンには約51個、中型タンパク質であるミオグロビンには約153個、大型タンパク質であるヘモグロビンには約574個、巨大タンパク質であるチチン (Titin)には約34,350個のアミノ酸の組み合わせが含まれている。20種類のアミノ酸の組み合わせによって小型タンパク質であるインスリンが形成される確率は1/2051=4.4 x 10-67であり、巨大タンパク質であるチチンが形成される確率は1/2034350=10-44680≈0である。これもまた、タンパク質形成がランダムな過程で形成されることは不可能であることを示唆している。
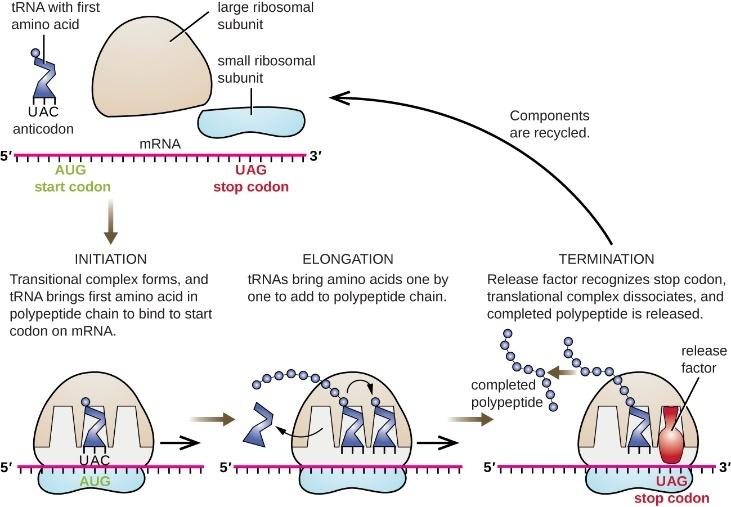
図3.2.タンパク質形成過程
アミノ酸からペプチド結合が形成される過程は多くのエネルギーを必要とするが、原始大気中でこの結合を促進する一貫した十分なエネルギー源が存在したかどうかという疑問も提起される。雷、紫外線、放射線、火山熱などの様々なエネルギー源が提案されたが、これらのエネルギー源が一貫してペプチド結合形成を促進できる効率と信頼性は議論の余地が多い。なぜなら、原始地球の環境は、極端な温度、pHレベル、環境変化など、過酷で変動が激しいものであった可能性が高いからである。このような条件は、むしろペプチド結合形成の繊細なプロセスを妨げ、形成されたペプチドの安定性を阻害する可能性がある。また、ペプチドは水溶性環境で加水分解されやすく、ペプチドが形成されるよりも早く分解される可能性がある。
iv. DNAの形成
DNAヌクレオチドは、RNAと同様に3つの部分で構成されている:窒素塩基(グアニン、アデニン、シトシン、チミン)、デオキシリボース糖、そしてリン酸基である。これらの自発的な形成は、RNAで見たようにほとんど不可能である。DNAの自発的な形成におけるもう一つの難関は、RNAとは異なり、追加的に二重らせん構造の塩基対を形成しなければならないことである。DNA二重らせんは、アデニンとチミン、シトシンとグアニンが特定の順序で配列されており、反対側の鎖には安全性のために補完的な配列が存在する。このように補完的な配列が完全に整列された構造であるため、案内テンプレートやメカニズムなしにランダムに作られることは不可能である。
自己複製はDNAの主要な機能の一つである。DNAの複製には、正確性と信頼性を確保するために複雑な酵素とタンパク質が必要だ。DNA複製に関与する主な酵素は、ヘリカーゼ、一本鎖結合(SSB)タンパク質、プライマーゼ、DNAポリメラーゼ、リボヌクレアーゼH(RNase H)、DNAリガーゼ、トポイソメラーゼなどがある。DNA複製に参加する酵素のアミノ酸総数は、通常数百個から数千個である。DNA複製にこれらの酵素がなければ、二重らせん複製およびエラー修正ができない。ところで、この酵素のいずれかが偶然に生成される確率は事実上0である。例えば、RNase Hが偶然に生成される確率は20-155≈2.2x10-202≈ 0で、現実的に発生する確率は全くない。
DNAは長い一本鎖で存在するのではなく、染色体の中で非常に複雑に折り畳まれたヌクレオソームの形で存在する。DNAが折り畳まれる過程をDNA凝縮過程(DNA condensation process)というが、この過程は多くのタンパク質の助けを借りて複数の段階の圧縮と折り畳み過程を経て行われる。二重らせん構造を持つDNAはヒストンタンパク質と結合してヌクレオソームを形成する。ヌクレオソームは、ヒストンタンパク質のコア(オクタマー:H2A、H2B、H3、H4それぞれ2個ずつ)と、これを約1.65回包んだ147個の塩基対で構成されたDNAで構成されている。その構造は、まるでビーズ(ヌクレオソーム)が紐(リンクDNA)にぶら下がっているように繋がっている。このヌクレオソームはさらに巻かれて直径30nmの繊維の形を形成し、螺旋状に配列されたり、ジグザグ状に配列されたりしている。30nmの繊維構造は、ループドメインと呼ばれるより大きな構造に折り畳まれる。このループドメインは、特定のタンパク質によって核内の特定の位置に固定され、遺伝子発現やその他の機能的な活動を調節する役割を果たす。ループドメインはさらに複雑に折り畳まれ、圧縮されて染色体を形成し、染色体は細胞分裂時にDNAを娘細胞に正確に伝達するために最も高いレベルで圧縮される。この段階で、コヒーシンとコンデンシンのようなタンパク質複合体が染色構造を安定化させ、正しく分離するのを助ける。
以上のように、DNAの形成と複製の過程は非常に複雑であり、様々な生化学的法則と複数の酵素の精巧な協力が必要不可欠である。このような複雑な過程に対する進化論的な説明は明確ではなく、単にDNAがRNAから転換されたとだけ主張されている。しかし、この主張が成立するためには、RNAがどのように形成されたのか、RNAからDNAの二重らせん構造がどのようなメカニズムで形成されたのか、そしてDNAの複製に不可欠な様々な酵素がどのような過程で生まれたのかなどを具体的に説明しなければならない。このような点を考慮すると、DNAは単純なランダムな過程の産物ではなく、精巧に計画された知的設計の産物であることを強く示唆している。
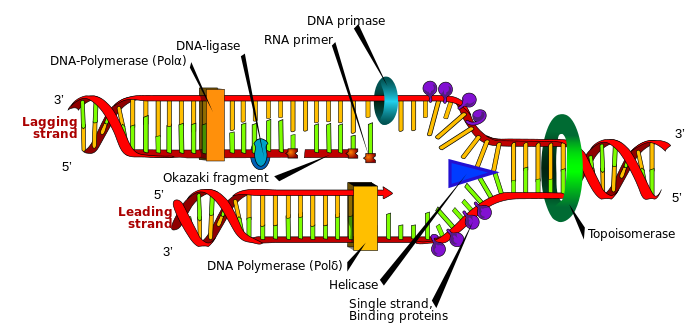
図3.3.DNA複製過程
v. 原核細胞の形成
議論を続けるために、RNA、タンパク質、DNAが偶然に形成されたと仮定すると、生命体に向けて次に越えなければならない山は、細胞の形成である。細胞には大きく分けて2つのタイプがある:原核細胞と真核細胞である。バクテリアや古細菌などの生物に見られる原核細胞は単純で、核がない。遺伝物質は、細胞質に自由に浮遊する単一の円形のDNA分子に含まれており、細胞小器官もない。科学者たちは、原始細胞が自然選択、突然変異、環境適応などの段階的な過程を経て原核細胞に進化したと主張している。しかし、原始細胞の存在は確認されたことがなく、原核細胞が地球上に最初に現れた細胞であったと考えられている。化石の記録によると、原核細胞は約35億年から38億年前に地球に現れた。すべての細胞は細胞膜にいるため、原始中で細胞が自然に形成された場合、その最初の過程はこの細胞膜の形成であったと推測される。では、細胞膜がランダムな過程を経て自然に形成されることができるのか調べてみよう。
- 細胞膜形成
細胞膜は単純な構造と考えられているが、実際には脂質(リン脂質、コレステロール、糖脂質)、タンパク質、そして炭水化物で構成される複雑でダイナミックな構造である。リン脂質は細胞膜の二重層構造を形成し、コレステロールは流動性を調節し、糖脂質は細胞認識に寄与する。タンパク質は内在性タンパク質と周辺部タンパク質に分けられ、運搬、信号伝達、構造的支持などの役割を果たす。炭水化物は細胞認識とコミュニケーションに重要な役割を果たす。
原始地球で機能性膜(functional membrane)に必要な特定の両親媒性(amphiphilic)脂質分子が形成されるためには、脂肪酸、グリセロール、そしてリン酸基が必要である。しかし、ダイナミックな原始地球の環境では、このような分子の正確な組み合わせが自発的に形成されることは難しい。さらに、前述したように、リン酸基の自発的な形成は不可能であり、このリン酸基がなければ細胞膜を形成することができない。
しかし、どうにかして両親媒性脂質分子が形成されたとしても、細胞環境をカプセル化して保護できる安定した半透過性の二重層が形成されなければならない。ところで、この二重層を形成するためには、十分な濃度の両親媒性分子(例えば、リン脂質)、中性に近いpH、膜の安全性を高めるカルシウムやマグネシウムのようなイオンの存在、高くも低くもない適切な温度、二重層形成に必要な水溶性環境、紫外線、熱、電気的放電などのエネルギー源、そして触媒表面の存在など、特定の条件が必要である。しかし、初期の地球の条件は、極端な温度、変動的なpH状態、強い放射線などで過酷で変動が激しく、このような環境で二重層膜が形成されたとしても、安定性を維持することは難しく、容易に破壊されただろう。
仮に、原核細胞の大きさを細菌細胞と同じ1マイクロメートル程度と仮定すると、その表面積は3x10-12m2となる。単一のリン脂質分子の大きさは約5x10-19m2なので、二重層を形成するために必要なリン脂質の数は約1,000万個となる。この1,000万個のリン脂質が並んで整列するのも大変だが、何らかの形の案内や方向性なしに自ら丸く曲がって3次元の閉鎖された細胞膜を作った可能性はほとんどないだろう。
細胞膜は必須栄養素と分子を選択的に通過させながら有害物質を遮断する機能を持たなければならない。このような選択的透過性は、複雑なタンパク質とチャネルの存在を必要とするが、これらがランダムな過程で形成され、勝手に細胞膜に統合される可能性は極めて低い。このように、細胞形成の第一段階である細胞膜が偶然に形成される確率が低いのであれば、生命体に向けた次の段階に進むことはできない。また、細胞膜がどのように自然に形成されたとしても、細胞膜内部を構成する細胞質、リボソーム、プラスミドなどがどのように形成されたかについての進化論的な説明が不足している。そして、これらがどのように偶然に形成されたとしても、どのようなメカニズムを通じて細胞膜の中にカプセル化されたのかについての追加的な説明も必要である。
vi. 真核細胞の形成
真核細胞の起源について広く受け入れられている説は、細胞内共生説(endosymbiotic theory)である。共生説は、真核細胞が原始的な原核細胞間の共生関係を通じて起源したことを示唆する。この過程は、特定の原核細胞(動物細胞の場合はミトコンドリア、植物細胞の場合は葉緑体)が祖先宿主細胞(ancestral host cell)によって吸収され、相互に有益な関係を形成し、最終的に複雑な真核細胞に発展したという。もっともらしいシナリオに聞こえるが、この理論が成立するためには、祖先宿主細胞と原核細胞がすでに存在していなければならない。祖先宿主細胞としては古細菌(archaea)を主張するが、問題は古細菌で原核細胞を細胞内に引き込む内包作用(endocytosis)が全く観察されなかったという点、そして古細菌の細胞膜はエーテル結合で構成されているが、真核細胞の細胞膜はエステル結合で構成されており、構造的に大きな違いがあるという点である。
先ほど、原核細胞が自然に生成されにくいと述べたが、同じ原核細胞に該当するミトコンドリアも自然に生成されることは 難しいと予想される。ミトコンドリアは、酸化的リン酸化を通じてATPを生成する細胞の発電所として機能する複雑な細胞小器官である。ミトコンドリアは外膜、膜間空間、内膜および基質(酵素、DNA、リボソーム、代謝物)などで構成されている。細胞膜と同様に、外膜はリン脂質とタンパク質の混合物からなるリン脂質二重層で構成されている。我々は先ほど、細胞膜がランダムな過程で形成されにくいことを明らかにした。また、外膜より複雑なDNAの自然形成も不可能なので、DNAを含むミトコンドリアが自然に形成される可能性は低い。
真核細胞における細胞核の起源は、さらに説明できない。細胞核は遺伝物質を保護し、細胞の代謝過程を効率的に調節する最も重要な細胞小器官である。細胞核は核膜に積み重なっている。進化論では、細胞核膜の起源について、膜内包仮説、ウイルス起源仮説、遺伝子転移仮説などを提示している。膜内包仮説は、原核細胞の細胞膜が内側に折りたたまれて核膜を形成したという仮説であり、ウイルス起源仮説は、ウイルスが原始真核細胞と相互作用をしながら遺伝物質を保護する核膜構造を形成したという仮説であり、遺伝子転移仮説は、異なる原核生物間の遺伝子転移及び混合が起こり、複雑な遺伝体が形成され、これを保護するための細胞膜が形成されたという仮説である。しかし、これらの仮説は次のような理由で妥当性がない。膜内包仮説の場合、細胞膜は単一のリン脂質二重層で構成されているが、核膜は二つのリン脂質二重層(外膜と内膜)で構成されており、核膜には核孔複合体という特殊な構造があるが、細胞膜にはなく、細胞膜と核膜に存在するタンパク質の種類と配置が異なるからである。ウイルス起源仮説の場合、通常、ウイルスは細胞の遺伝物質を自分の複製によって宿主細胞に利用するが、むしろ保護するために核膜を形成したという説明は説得力がない。遺伝子転移仮説は、転移自体が細胞内の遺伝子の多様性を増加させることができるが、核膜のような複雑な構造の形成を説明するには不十分であり、そのメカニズムも不明である。
核膜の中には核小体と染色体が存在するが、これらの起源についても納得できる説明がない。核小体と染色体には生命体の遺伝情報が含まれており、RNA、タンパク質、DNA、細胞小器官、生命体の組織および器官を形成する設計図が含まれている。生命体が形成される前の真核細胞の段階で、生命体を構成する設計図がすでに核の中に入っているという事実は、進化論的には絶対に説明できないし、むしろ生命体の知的設計を示す明確な証拠となる。
共生説が支持されるためには、原核細胞と真核細胞間の移行形態を示す化石的証拠が必要であるが、そのような化石は発見されていない。また、ミトコンドリアは自身のDNAと独立して複製する能力など、いくつかの自律性を維持している。もしミトコンドリアが真核細胞に統合されたのであれば、数十億年の進化の後、ミトコンドリアの遺伝子が宿主遺伝子に完全に統合されていないという疑問も提起される。さらに、ミトコンドリアを構成するDNA、酵素、リボソームなども自然に形成されることは不可能である。
vii. 細胞小器官の局在化
細胞内には複数の細胞小器官が存在する。細胞小器官の局在化(organelle localization)は、細胞内で細胞小器官が特定の位置に配置される過程である。核は細胞中央、ミトコンドリアは細胞周辺部、小胞体は核付近、小胞体付近、そしてリソソームは細胞質に位置する。細胞小器官が適切な位置にある時、細胞の機能が最適化され、細胞の構造的統合性と機能的調和がよく維持される。細胞小器官は自分で考えることができないはずなのに、どのようにしてそのような最適な場所に自力で行くのだろうか?細胞小器官の局在化過程を詳しく見ると、それが非常に複雑であることが分かり、このようなメカニズムが偶然に形成されたのではなく、精巧な設計によって調整され、進行された過程であることを確認することができる。
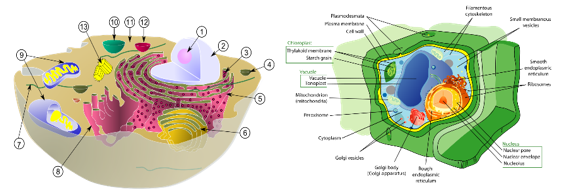
図3.4.動物細胞と植物細胞の構造
細胞小器官の局在化は、細胞骨格、微小管、モータータンパク質、膜輸送、アンカータンパク質と足場タンパク質、動的調節、細胞間コミュニケーションなどの複雑な相互作用によって行われる。
細胞骨格は細胞の構造的支持を提供し、微小管、アクチンフィラメント、中間フィラメントの3つの主要なタイプのフィラメントで構成されている。各フィラメントは、細胞小器官の局在化において特定の役割を担っている。
微小管は、チューブリンタンパク質で構成された長くて中空の管であり、微小管形成中心(中心体)から細胞周辺につながるネットワークを形成する。微小管は、中心体の近くに位置するゴルジ体と細胞の様々な場所に分布しているミトコンドリアのような小器官をキネシンとダイニンのようなモータータンパク質の助けを借りて細胞内の適切な場所に移動させる道路網の役割をする。キネシンは細胞小器官を微小管のプラス端、一般的に細胞周辺に移動させ、ダイニンはマイナス端、一般的に細胞中心側に移動させる。
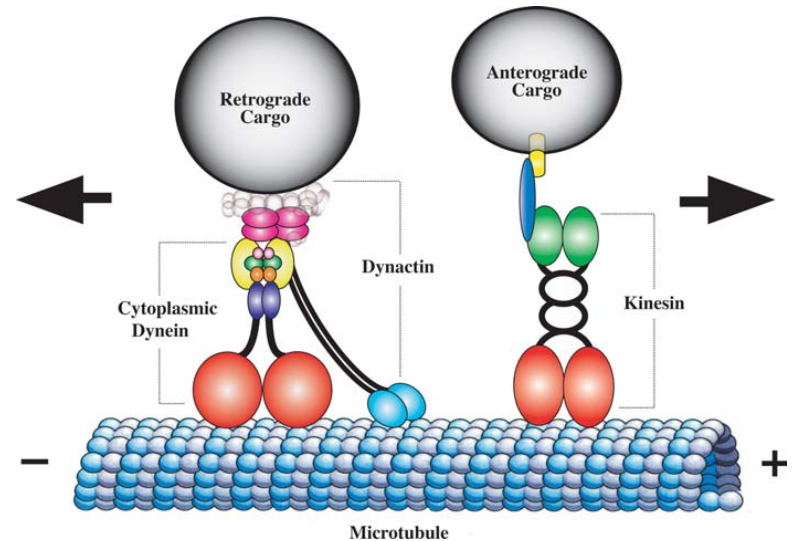
図3.5.微小管とモータータンパク質
マイクロフィラメントとも呼ばれるアクチンフィラメントは、アクチンタンパク質で作られた細くて柔軟な繊維である。アクチンフィラメントは原形質膜のすぐ下に集中しており、細胞質全体に密なネットワークを形成している。アクチンフィラメントは、細胞小器官と栄養素を細胞全体に分配する過程である細胞質ストリーミングの役割を促進する。ミオシンモータータンパク質はアクチンフィラメントと相互作用し、アクチンネットワークに沿って小胞体、エンドソーム、その他の小器官を運ぶ。アクチンフィラメントは、細胞の形状を維持し、細胞の移動に関与し、小器官の位置に間接的に影響を与える。
中間フィラメントは、細胞の種類に応じて様々なタンパク質(ケラチン、ビメンチン、ラミンなど)で構成されたロープのような繊維で、細胞の機械的強度と構造的支持力を提供する。中間フィラメントは、核などの小器官を細胞質内に固定し、その位置を安定させるのに役立つ。また、細胞骨格の全体的な完全性を維持し、微小管やアクチンフィラメントなどの他の構成要素が細胞小器官の局在化に効果的に機能できるようにする。
様々な種類の細胞骨格フィラメントは、多くの場合、細胞小器官を正確に配置するために協働する。例えば、微小管とアクチンフィラメントは、小胞(vesicle)と小器官の適切な分布と移動を保証するために協調する。細胞骨格は非常にダイナミックであり、細胞のニーズに応じて継続的にリモデリングされる。この柔軟性のおかげで、細胞シグナルや環境の変化に応じて細胞小器官を迅速に再配置することができる。
膜輸送は、細胞内で小胞を介してタンパク質、脂質、その他の分子を細胞内の正確な目的地に輸送するプロセスである。クラトリンなどの特定のタンパク質が関与し、寄付された膜の特定の部位で小胞が形成され、輸送する貨物分子を小胞に包む。形成された小胞は、モータータンパク質の助けを借りて、細胞骨格内のアクチンフィラメントと微小管に沿って移動する。そうして小胞が目的地に到着すると、目的地の膜と融合して内容物を放出する。この過程で、SNAREタンパク質のような特殊なタンパク質が小胞と目的地の膜を認識し、結合を助ける。膜輸送に関与する主要な細胞小器官には、小胞体、ゴルジ体、エンドソーム、リソソームなどの様々なタイプの小胞体がある。このプロセスは、細胞組織を維持し、小器官間のコミュニケーションを促進し、細胞が内部および外部信号に効率的に反応できるようにする。
シグナル伝達経路は、空間的な手がかりを提供する化学的シグナル伝達を通じて、細胞小器官が適切な位置に移動できるようにする。細胞質表面と細胞質内の受容体は、このプロセスを促進するためにシグナル分子と相互作用する。例えば、Rabタンパク質のような小さなGTPaseは、特定のエフェクタータンパク質と相互作用し、小胞の移動と小器官の位置を制御する重要な調節因子である。これらのシグナル伝達経路は、変化する細胞の要求と環境条件に応じて細胞プロセスが調整され、細胞小器官が動的に配置されるようにする。
アンカータンパク質とスカフォールドは、細胞小器官が細胞内に正確に位置するようにすることで、細胞の位置決定に重要な役割を果たす。アンカータンパク質は細胞質内の特定の部位に小器官を接続し、小器官を安定化し、小器官の移動を防止する。例えば、ミトコンドリアは特定の固定メカニズムを通じて小胞体に固定され、効率的なエネルギー伝達と代謝調整を促進することができる。足場タンパク質は、細胞小器官を所定の位置に固定する複合体を形成し、細胞全体の組織を維持することで構造的なサポートを提供する。これらのタンパク質は、細胞小器官の適切な配置を可能にする動的な枠組みを作り、細胞機能が効果的かつ効率的に実行されるようにする。
細胞位置の動的調整とは、細胞内の小器官の位置が連続的かつ反応的に調整整列する過程を指す。このような調整は、細胞の 機能と適応性を維持するために非常に重要である。有糸分裂などの細胞周期の様々な段階で、核やミトコンドリアなどの細胞小器官は、適切な細胞分裂のために位置を動的に変化させる。また、栄養素の可用性やストレス条件などの環境刺激に反応して、細胞小器官はその機能が最も必要とされる領域に再配置されることがある。このような動的再配置は、細胞骨格と運動タンパク質によって促進され、細胞が恒常性を維持し、変化する内部および外部条件に効率的に対応できるようにする。
細胞間コミュニケーションとは、細胞内の様々な小器官が互いに信号を送受信し、物質を交換しながら協力する過程を指す。細胞はこのような通信を通じて生理学的機能を調節し、恒常性を維持し、外部刺激に効果的に対応することができる。細胞間のコミュニケーションは、直接接触部位と小胞体輸送を通じて行われ、細胞の機能を調整し、恒常性を維持するために不可欠である。ミトコンドリアと小胞体間のミトコンドリア関連膜(MAMs)などの接触部位は、脂質、カルシウム、その他の分子の移動を促進し、細胞小器官間の同期化された活動を保証する。小胞輸送には小胞の分化と融合が含まれ、小胞は細胞小器官間でタンパク質と脂質を輸送し、細胞小器官の機能的統合を維持する。効果的な細胞間コミュニケーションは、代謝、シグナル伝達、ストレス反応などのプロセスに最も重要である。
以上のように、細胞小器官の局在化メカニズムは高度に複雑で精巧に組織化されており、明確な目的と方向性を持っている。進化論では、このような精巧なシステムがランダムな突然変異と自然選択によって徐々に形成されたと主張する。この主張が認められるためには、まず、モータータンパク質、アンカータンパク質、足場タンパク質のような重要な構成要素がランダムな突然変異だけで形成されるのかについて具体的な説明が必要である。また、膜輸送、動的調節、細胞間コミュニケーションなどの必須メカニズムがどのように確立されたかについての実質的な証拠が提示されなければならない。特に、細胞小器官の局在化メカニズムが段階的に進化したという主張を裏付けることができる中間段階の化石や生物学的証拠も提示されるべきである。さらに、細胞小器官の局在化に関与するタンパク質とメカニズムのいずれかが欠けても機能を遂行することができないという点で、これらの複雑な要素がどのように完璧な調和を成して同時に存在するようになったかについての説明が求められる。しかし、進化論ではこの ような核心的な事実を全く説明できない。
細胞小器官の局在化過程は、まるでアマゾンで商品を注文すると正確な住所に配送される過程と似ている。アマゾンの物流輸送体系が偶然に形成されたのではなく、精巧な計画と綿密な管理によって構築されたように、細胞小器官の局在化メカニズムもランダムな過程の結果ではなく、精巧で緻密な設計によって形成されたことを明確に示している。このような秩序ある配置と機能の調和は、自然発生的な偶然の産物ではなく、高度な知性の存在を示唆していると言えるだろう。
viii. 細胞分化
細胞分化とは、非特化細胞から特定の機能を持つ特化細胞に分化する過程を指す。細胞分化は生命体の組織および器官形成に必須であり、幹細胞から始まり、遺伝子発現調節、細胞外シグナル、細胞間相互作用など様々なメカニズムによって細胞分化が調節される。
遺伝子発現調節は、転写因子タンパク質が特定の遺伝子の発現を活性化したり、抑制したりして細胞の運命を決定する。例えば、MyoDは筋肉細胞に、Sox2は神経細胞に分化させる。この過程には、DNA配列の変化なしに遺伝子発現を調節するDNAメチル化、ヒストン修飾、非暗号化RNAの過程が入るが、これをエピジェネティック調節過程と呼ぶ。DNAメチル化は、DNAのシトシン塩基にメチル基を追加する過程で、通常、遺伝子発現を抑制する。ヒストンタンパク質の修正(アセチル化またはメチル化)は、染色質の構造を変化させて遺伝子アクセス性を調節し、ヒストンアセチル化は遺伝子発現を活性化し、メチル化は遺伝子発現を抑制する。非暗号化RNA(ncRNA)はタンパク質に翻訳されないが、遺伝子発現の調節と細胞分化過程で重要な役割を果たす。例えば、miRNAはmRNAと結合したり、分解したりして、遺伝子発現を調節する。siRNAもmRNAと結合して分解することで特定の遺伝子発現を抑制する。長鎖非翻訳RNA(lncRNA)は様々なメカニズムを通じて遺伝子発現を調節することができ、染色体リモデリング、転写調節、翻訳調節などに関与する。snRNAとsnRNPはスプライシング過程でイントロンを除去し、エクソンを連結する役割を果たし、snoRNAはrRNAのメチル化を促進する。
細胞外シグナルは、細胞外から分泌される物質や因子で様々な形態が存在し、特定の細胞の表面受容体によって認識され、細胞内MAPK/ERK、PI3K/Akt、Wnt/β- Catenin経路などのシグナル伝達経路を通じて細胞内部に伝達される。外部シグナルの主な種類には、成長因子、モルフォゲン、ホルモン、サイトカインなどがある。成長因子は細胞の成長、分化、生存を促進するタンパク質であり、表皮細胞はEGF、繊維芽細胞はFGF、血管内皮細胞はVEGFなどがある。モルフォゲンは濃度勾配によって細胞の運命を決定するシグナル分子で、神経管の発達を調節するShh、骨と軟骨形成を誘導するBMPsなどがある。ホルモンはよく知られているように、インスリン、エストロゲン、テストステロンなどがあり、血流を通じて標的細胞に移動し、特定の生理学的反応を誘導するシグナル分子である。サイトカインは免疫反応を調節するタンパク質で、免疫細胞間の信号を伝達するインターロイキン、ウイルス感染に対する免疫反応を促進するインターフェロン、炎症反応と細胞死を誘導するTNFなどがある。
細胞間相互作用は、細胞が互いに信号を交換して細胞の運命を決定し、組織の形成と機能を調節するプロセスである。このような相互作用は、細胞の表面にある受容体とリガンド間の結合を通じて行われ、特定のシグナル伝達経路を活性化して細胞分化を誘導する。細胞間相互作用のメカニズムには、Notchシグナル伝達経路、ギャップ結合(gap junctions)、カドヘリンとインテグリンが介在する接触依存的なシグナル伝達、近くの細胞にシグナルを伝達するパラクリンシグナル伝達、モルフォゲン濃度勾配を通じたシグナル伝達などがある。
以上で見たように細胞の分化過程は、遺伝子発現、外部シグナル、細胞間相互作用などの様々な複雑で精巧なメカニズムが互いにうまく協調して進行していることが分かる。遺伝子発現調節の場合、細胞分化の特定の段階と場所で正しいシグナルを受けた時のみ、特定の遺伝子が活性化される。この過程で、数百の転写因子、エンハンサー、リプレッサーおよびその他の調節要素が相互作用する複雑なネットワークを通じて遺伝子発現を調節する。細胞外シグナルは、特定のリガンドが特定の受容体に結合し、多段階のシグナル伝達経路を活性化し、細胞内で特定の反応を誘導する。細胞間の相互作用も、特定の細胞が特定の位置で特定の役割を果たすように誘導され、正と負のフィードバックによって調節される。
進化論では、細胞分化も突然変異と自然選択によって徐々に形成されたと主張するが、これを裏付ける具体的な証拠が不足している。まず、細胞分化は単純な生化学的反応ではなく、明確な目的と方向性を持っているが、突然変異と自然選択にはこのような方向性がない。進化論では、単細胞生物が徐々に多細胞生物に発展しながら細胞分化が行われたと主張するが、単細胞から多細胞への転換過程、そして組織と器官が形成される中間段階を証明する実質的な化石的・生物学的証拠は存在しない。現在発見された多細胞生物の化石はすでに完全な形で分化した状態であり、段階的に複雑性が増加する過程を示していない。また、細胞分化は、遺伝子発現、外部シグナル、細胞間の相互作用が精密に調整された状態で同時に作動しなければならない過程であり、どれか一つでも欠けていたり、進化の途中であったりすると、作動しない。このような様々な事実は細胞分化がランダムな突然変異と自然選択の産物ではなく、精緻な知的設計の結果であることを明らかに示している。
ix. 組織と機関の形成
生命体への細胞分化の次の段階は、組織と器官の形成である。細胞の組織形成は、幹細胞が細胞間相互作用、細胞移動、細胞外基質の複合的な作用により、細胞が集まって特定の機能を遂行する組織を形成する。
細胞間相互作用は、細胞が互いに信号を交換し、物理的に結合して組織を形成する過程である。相互作用シグナル伝達は、Notchシグナル伝達経路とWntシグナル伝達経路を使用する。結合に関与する接着タンパク質分子として、カドヘリンは細胞間の結合を媒介し、組織の構造的統合性を維持し、インテグリンは細胞と細胞外基質間の結合を媒介し、組織の安定性を提供する。
細胞移動は、細胞が特定の位置に移動して組織を形成する過程である。化学信号に沿って細胞が移動し(ケモタキシス)、微細繊維(アクチンフィラメント)のような細胞骨格要素が再構成されて細胞移動を促進する。細胞移動は胚の発達、創傷治癒、免疫反応などに重要な役割を果たす。
細胞外マトリックス(ECM)は、細胞を取り囲んで支える複雑なネットワークである。構成要素には、組織の構造的強度を提供するコラーゲン、組織の弾力性を提供するエラスチン、ECMの水分含有量と圧縮抵抗性を調節するプロテオグリカンなどがある。ECMは、細胞が組織内で安定的に位置するように機械的なサポートを提供し、細胞と相互作用して細胞の分化、移動、生存を調節する信号を伝達する。
このような過程を通じて生成された組織の主なタイプは、皮膚や腸粘膜のような保護、吸収、分泌機能を実行する上皮組織、骨、軟骨、脂肪のような支持と結合機能を実行する結合組織、骨格筋、心筋、平滑筋のような収縮を通じて運動が可能な筋肉組織、脳、脊髄、神経のような信号伝達機能を実行する神経組織などである。これらの組織が相互補完して上位組織を形成するが、例えば、上皮組織と結合組織が調和して皮膚組織を形成するのである。
このように、組織形成には非常に複雑な遺伝子調節ネットワークが関与しており、多数の遺伝子とその発現を精巧に調節する様々な転写因子、エピジェネティックメカニズムなどが含まれる。また、信号伝達、接着、移動などの細胞間相互作用が調和して初めて組織が正しく形成される。このような精巧な遺伝子発現調節機能と細胞間の相互作用が、ランダムな突然変異と自然選択の段階的な変化によって形成されたと見ることは不可能である。なぜなら、組織形成の中間形態が十分な選択的利点を提供したならば、自然選択によって保存されただろうが、機能的な中間形態の不在は、組織形成が段階的な進化の結果であると見ることができないからである。何よりも、組織形成のような複雑な構造が徐々に進化したことを証明する中間段階の化石の証拠がない。そして、組織形成には様々な細胞タイプと組織が相互に依存して働く必要がある。例えば、筋肉組織は筋肉細胞だけでなく、血管や神経組織も必要である。しかし、これらの相互依存的なシステムが同時に進化する可能性は非常に低い。
組織が形成されたら、組織から器官が形成されなければならない。この過程は、異なるタイプの組織が相互作用して複雑な構造と機能を持つ器官を作る過程である。例えば、筋肉組織、結合組織、神経組織などが結合して心臓を作るのである。臓器が作られると、臓器内の細胞と組織が特定の機能を遂行するようにさらに分化して専門化される機能的分化が起こらなければならない。このように臓器が形成されると、発生学と細胞生物学の精巧な調節メカニズムによって臓器が決まった位置に位置するようになる。
x. 多細胞生物の形成
個々の器官が形成されると、それらは単に存在するだけでは不十分である。すべての臓器は体内の特定の位置に配置され、互いに緊密に統合されて初めて、一つの完全でよく機能する生命体を形成することができる。このような統合は、ランダムな配置ではなく、非常に精密に調整された位置選択と接続の過程を必要とする。各臓器は、体内の他の臓器および器官系(例えば、循環系、呼吸系、消化系、神経系など)と機能的に適切に相互作用できる位置に正確に配置されなければならない。例えば、心臓と血管で構成される循環器系は、酸素を供給する呼吸器系、栄養素を伝達する消化器系と物理的に隣接している必要があり、同時に機能的にも適切に接続されている必要があります。もしこのような接続が不正確または不十分であれば、酸素と栄養分の伝達という本質的な機能が適切に行われず、生命維持が不可能になる。この複雑な配置と統合の過程で、各臓器を構成する組織と器官内の細胞は絶えずそれぞれの固有の役割に合わせて専門化され、適応していく。このような過程を機能分化(functional differentiation)という。機能分化を通じて、生命体の各部分は自分に与えられた特定の機能をより効果的に遂行できるようになり、生物全体は一つの調和のとれたシステムとして機能できるようになる。特に、複数の器官と器官系間には様々な生化学的信号と物質交換が行われるが、この相互作用は単純な接続を超えた高度に複雑な体系を形成する。この複雑なシステムは、一つでも欠落するか、未完成の部分が存在すると、生命体全体が正常に機能できなくなる。言い換えれば、生命体は、すべての主要な器官と器官系が最初から完全な形で存在し、互いに正確に接続されなければ、完全な機能を発揮することができない。このような複雑で精密な相互依存性は、自然淘汰、突然変異、または緩やかな進化といった説明だけでは理解できない構造を示している。 なぜなら、一つの器官が部分的にしか発達していない、または一つの器官系が未完成の状態で存在する場合、生命体が、生存はおろか、基本的な機能すら実行できないからだ。このように、生命体を構成する器官と器官系は『all or nothing』システムを形成しており、これは最初から完成された状態で一緒に存在してこそ可能な非常に精巧な設計と言える。
ここまで見てきたように、生命体を構成する主要要素が自然な過程だけで偶然に発生する可能性は、いくつかの単純なアミノ酸を除いて、事実上全くないことが明らかになった。むしろ、実験的結果と理論的分析は、複雑な生命体の構成要素がランダムな化学反応では決して自然に形成されることができないという事実をより強く支持している。また、アミノ酸が結合してタンパク質を形成し、タンパク質と核酸が精巧に組み立てられてDNAとRNAを形成し、さらに細胞、組織、器官に至る一連の過程には、単純な化学反応を超えた明確な方向性と設計意図が存在することが分かる。この一貫した流れと目的性は、ランダムな突然変異と自然選択という進化論的説明では決して説明できない特徴である。むしろ、これは生命体が最初から高度な知性と計画によって意図的に設計されたことを強く示唆している。さて、次の節では、このような観点から、進化論が果たして生命体の起源を合理的に説明することができるのかどうかを見てみよう。
b. 進化論は生命の起源を説明できるのか?
進化論ではこの過程を説明するために様々な理論を提示してきたが、その主な理論は自然選択、突然変異、遺伝的浮動(genetic drift)、水平的遺伝子移動(horizontal gene transfer、HGT)などがある。各理論について簡単に説明しよう。
自然選択とは、有利な形質を持つ個体がより正常に生存・繁殖し、何世代にもわたってその形質が個体群にますます多く現れる過程である。自然選択は既存の生命体の変異に適用され、生命体の基本構成要素(アミノ酸、RNA、タンパク質、DNA)および構造(細胞、組織、器官、多細胞生命体)形成には適用されないため、生命体の初期発生を説明することはできない。
突然変異は、遺伝子の塩基配列にランダムに起こる変化を指し、自然選択と同様に、生命体の初期発生と高度に組織化された複雑な構造を説明できない。さらに重要な問題は、ほとんどの突然変異が有害または中立的なものであることである。有益な突然変異は非常にまれであるため、これらの突然変異が蓄積して有意な変化を引き起こすことは困難である。例えば、Vesicular Stomatitis Virusの研究で変異の結果を見ると、約40%が生存に致命的な方向(fitness=0)、約31%が致命的ではないが有害な方向(0
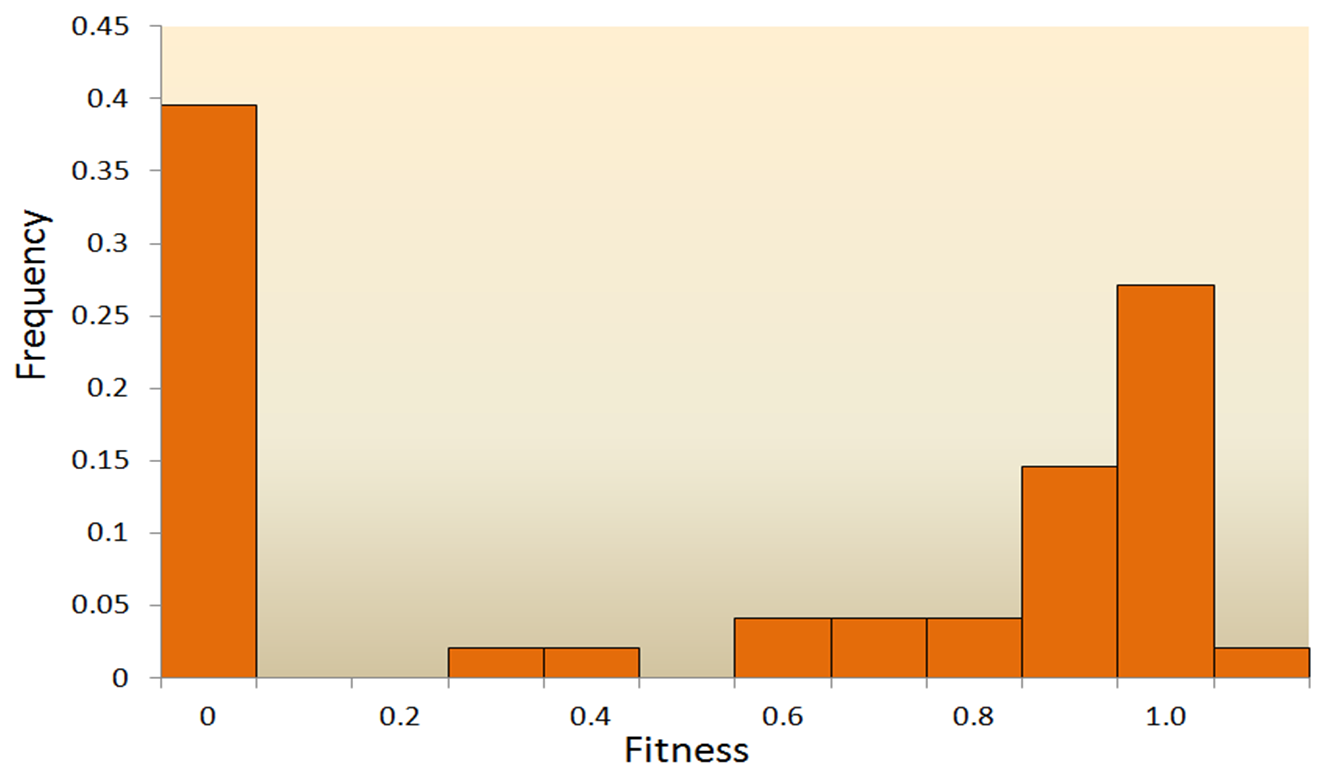
図3.6.突然変異の適応度(fitness)の分布
遺伝的浮動は、遺伝子頻度のランダムな変化を通じて集団の遺伝的構成を変化させるという理論である。遺伝的浮動は小さな集団でより顕著であり、ほとんどの変化が起こる大きな集団では影響が少ない。また、高度に組織化された構造やシステムの発達を説明するための方向性を欠いている。さらに、遺伝的浮動は新しい情報や機能を生み出さないため、新しい特性の出現や複雑な生物学的特徴の起源を説明することができない。
遺伝子の水平伝播(HGT)は主に原核生物に限られた役割を果たすため、多細胞生物の複雑な特性を説明することはできない。HGTは外来遺伝子を宿主のゲノムに組み込むため、遺伝的不安定性をもたらし、有害な突然変異につながる可能性がある。また、HGTは新しい遺伝子の起源を説明するのではなく、既存の遺伝子の転移を説明するため、新しい特性の出現には対応できない。次の表は、生命の起源に対する進化論の適用可能性をまとめたものである。
進化論 | 生命の起源説明可能? | RNA、タンパク質、DNAの形成を説明可能? | 進化論ではなく遺伝的適応論か?* |
自然選択 | 不可能 | 不可能 | Yes |
突然変異 | 不可能 | 不可能 | Yes |
遺伝的浮動 | 不可能 | 不可能 | Yes |
HGT | 不可能 | 不可能 | 該当なし |
表3.2.生命の起源に対する進化論の適用範囲(*: 遺伝的適応論については次のセクションを参照)
上記の表から分かるように、進化論の主要理論は、地球生命体の起源だけでなく、RNA、タンパク質、DNAなどの生命体の基本構成要素がどのように形成されたかについての具体的なメカニズムを提示していない。特に、生命体の形成過程は高度に複雑で精巧な方向性を持っているにもかかわらず、方向性のないランダムな過程だけでこれを説明するには限界がある。それでも、ほとんどの生物学の教科書や『利己的遺伝子』のような進化論を擁護する書籍では、遺伝子と生命体が原始地球環境で化学的に偶然に生成されたという仮定を前提としている。しかし、実際には、徹底的に制御された実験室環境でも、たった数個のアミノ酸が合成されるのが精一杯であり、さらに、これらのアミノ酸がRNA、タンパク質、DNAに自然に形成される過程は、生化学的にも、確率的にも全く可能性のないことである。また、このような複雑で精巧な条件を要求する生命体が、汚染され混濁した深海熱水域で誕生したという、まったく受け入れがたい主張をする人もいる。
生命体は変化する環境に対して絶えず適応し、生存を維持していく。この適応能力はすでに生命体の遺伝子に記録されており、必要に応じて適切な遺伝子が発現されて環境に適応する。進化論は単に変化する環境に生命体が適応する姿を描写しているに過ぎず、進化論と呼ばれるよりは「遺伝的環境適応論」と呼ばれるべきである(次のセクション参照)。このような重要な限界にもかかわらず、進化論は過度に宣伝され、ほとんどの人は進化論が生命体の起源と様々な生命体の形成を説明する理論だと誤解している。
建物を建てるためには、設計図、建築材料、強固な基礎が必要である。生命体の形成をビル建設に例えると、進化論は設計図(方向性)、建築材料(RNA、タンパク質、DNA)、基礎(生命体の起源)なしでランダムにビル建設が行われるのと同じだ。しかし、ビルは設計図なしで建設されることはできず、よく設計された設計図通りに施工されなければビルが完成することはできない。ビル建設に設計者が存在するように、ビルとは比較にならないほど複雑な生命体形成にも設計者が必ず存在しなければならない。
生命体を設計し、創造されたのは、創造主である神様である。
c.ダーウィンの理論は進化論か?それとも遺伝的適応論なのか?
生物の進化は、同じ種間で変異が起こるMicroevolution(小進化)と、ある種が別の種に変わるMacroevolution(大進化)に分けることができる。進化論者たちは、自然選択や突然変異による小進化が蓄積されて大進化が起こったと主張する。そのためには、ある種から別の種に移行する中間種の存在が必然的に存在しなければならないが、どこを見ても中間種は存在しない。ダーウィンもこの事実を知っていて、種と種の間の中間種が存在せず、自分の理論を主張するのに困難があると告白している。《種の起源》第6章「進化論の難関」の最初の部分を見ると、こう書かれている:「もし種が微細な段階的な変化によって他の種からしたのであれば、なぜ私たちはどこでも無数の転移形態の中間種を見ないのか?」この事実はダーウィンのジレンマ(Darwin's dilemma)として知られており、自分の進化論が何か間違っていたことを認める告白である。
かつて約3億7,500万年前に存在したTiktaalikが魚と中間化石であるという主張があった。しかし、Niedzwiedzkiらが発表した2010年のNatureに掲載された論文を見ると、Tiktaalikより約1,800万年前の四足動物の足跡の化石を見ることができる。Tiktaalikより古い四足動物の足跡の存在は、Tiktaalikが魚と四足動物の中間種ではないことを示唆している。リング種(ring species)の概念で中間種の存在を説明しようとするが、無理な主張であることが分かる。なぜなら、リング種は種と種の間の中間種ではなく、同じ種内で生物学的交配が起こらない種だからである。様々な遷移段階の中間種の化石がないため、ダーウィンの理論は進化論と呼ぶことができず、変化する生存環境に適応する生命体の遺伝的適応論と呼ぶべきである。その主な理由は、次のミランコビッチサイクルと関係している。
- ミランコビッチサイクル
地球はほぼ円軌道を回っているが、約9万6千年周期で偏心率が変化する。偏心率の変化は気候変動に影響を及ぼし、氷期と間氷期の時期にも影響を与える。そして現在23.5度傾いている自転軸の傾きは、41,000年周期で21.5度から24.5度の間で変化する。この傾きの変化は、赤道と極地の間の太陽輻射分布に影響を与え、季節の強度を変化させ、長期的な気候パターンと氷河期の力学に重要な役割を果たす。地球の自転軸は洗車運動により、26,000年周期で軸の方向が徐々に変化する。洗車運動は季節の強度と時期を変化させ、地球全体の気候システムに影響を与える。
偏心率、自転軸の傾き、自転軸の洗車運動の変化が組み合わさった効果は、ミランコビッチサイクルと呼ばれ、この周期は長期的な世界的な気候変動を引き起こす。その良い例がサハラ砂漠の気候変動である。太陽放射が増加する期間中、サハラはより多くの降雨を経験し、湖や川がある緑豊かな風景に変わり、逆に太陽放射が減少すると乾燥した状態になり、今日のような広大な砂漠になる。
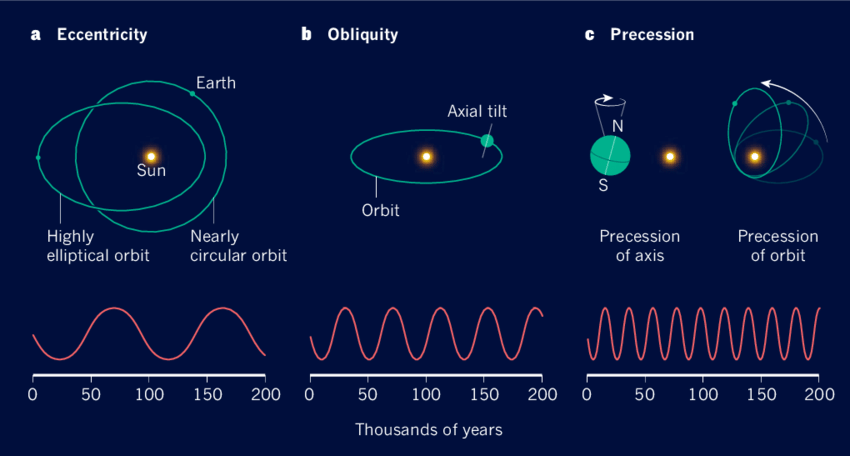
図3.7.ミランコビッチサイクルの構成要素
地球にこのような広域的な気候変動が発生すると、地球上のすべての生命体は遺伝的適応を通じて環境変化に身を合わせる。遺伝的適応は、生命体が絶滅することなく長い期間、地球上で生存できるようにする驚くべき生物学的メカニズムであり、創造主である神様が生命体の符号化、非符号化DNAの中に隠しておいた驚くべき環境適応力である。このような適応力のため、ミランコビッチサイクルを経験しても生命体が消滅することなく、今まで地球上でうまく生存してきた。進化論者は、このような適応力を「進化」という誤った名前で呼んでいる。しかし、正確には「遺伝子の環境適応論」または「遺伝的適応論(Theory of genetic adaptation)」と呼ぶべきである。では、次に遺伝的適応論のいくつかの例を挙げてみよう。
- 紫外線に対する遺伝的適応
気候変動により人間の皮膚が強い紫外線(UVB、UVA)にさらされると、複数のタンパク質とホルモンが関与する複雑なメカニズムを通じてメラニン生成が増加する。紫外線が皮膚細胞のDNAに損傷を起こすと、細胞のストレスと損傷に対する反応を調節する重要な調節因子であるp53タンパク質が活性化する。活性化されたp53タンパク質は転写因子として作用し、紫外線損傷に対する保護反応に関わる様々な遺伝子の発現を促進する。
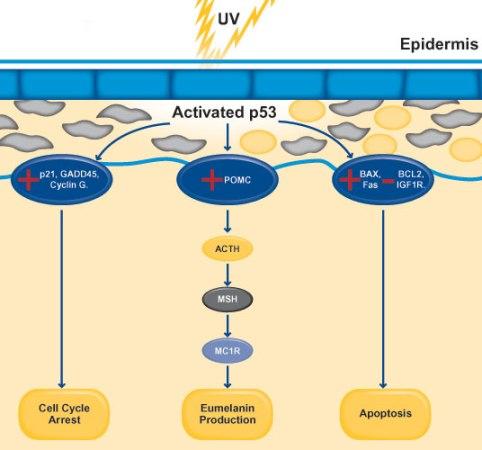
図3.8.遺伝的適応によるメラニン生成メカニズム
p53はPOMCと呼ばれるホルモン前駆体タンパク質遺伝子の発現を誘導し、POMCはACTHと呼ばれる副腎皮質刺激ホルモンを生成し、ACTHはMSHと呼ばれるメラニン細胞刺激ホルモンを生成する。生成されたMSHは、メラニン細胞表面のMC1Rと呼ばれるメラノコチン1受容体に結合する。MSHのMC1R結合は、メラニン細胞内部でシグナル伝達連鎖反応を引き起こし、メラニン合成に関わる遺伝子の発現を増加させる。
メラニン細胞はメラニンの生産を増加させ、生産されたメラニンはメラノソームに包まれ、皮膚の外層を構成する主要な細胞タイプである角質形成細胞に移動する。移動したメラニンは角質形成細胞の核の上に保護膜を形成し、紫外線を吸収して分散させ、皮膚細胞のDNAをさらなる紫外線ダメージから保護する。このような一連の複雑な過程は、生命体が変化する環境に適応する生物学的メカニズムの一つであり、遺伝的適応が比較的短い時間 内に起こることができる良い例である。
- 北極環境への遺伝的適応
北極圏付近に住むイヌイット族は、厳しい北極圏の環境で生き残るための遺伝的適応をしてきた。主な遺伝的適応には、脂肪酸脱飽和酵素(FADS)遺伝子群の変異が含まれ、海洋哺乳類の伝統的な高脂肪食からオメガ3およびオメガ6脂肪酸を代謝する能力を向上させる。また、カルニチンパルミトイルトランスフェラーゼI(CPT1A)遺伝子の遺伝的変化は、脂肪からのエネルギー生産を改善し、体温維持に不可欠な役割を果たす。このような適応は、高脂肪食にもかかわらず、心血管疾患のリスクを軽減する。さらに、褐色脂肪の活動を調節する遺伝子の適応は、熱産生を増加させ、イヌイットが極寒の中で熱を発生させ、体温を維持するのに役立っている。
このような遺伝的適応は、少なくとも20,000年前、イヌイットの祖先がロシアとアラスカの間のベーリング海峡周辺に住んでいたときから始まったようで、これは変化する環境に対する遺伝的適応のもう一つの良い例である。

図3.9.寒冷な環境に遺伝的に適応したイヌイット族
- ヒグマからホッキョクグマへの遺伝的適応
ヒグマからホッキョクグマへの移行は、環境変化によって誘発された遺伝的適応の別の例である。約40万年前、ヒグマの集団が北極圏で孤立し、異なる生存環境に直面した。時間が経つにつれて、過酷で氷に覆われた環境で有利な遺伝的変化が起こり、主な適応には、脂肪代謝に関連する遺伝子、例えばアポリポタンパク質B(APOB)遺伝子などの変化が含まれる。これにより、主食であるアザラシの高脂肪食を処理する能力が向上した。エンドセリン受容体B型(EDNRB)やAbsent in Melanoma 1(AIM1)などの遺伝子の適応は、白い毛皮の発達につながり、雪や氷の中でカモフラージュできるようになった。さらに、クマの骨格構造と手足の形態に影響を与える遺伝的変化は、北極海域での狩猟に重要な水泳能力を向上させた。
このような遺伝的適応は、ホッキョクグマが北極の資源を効率的に利用し、極端な寒さに耐え、ヒグマの祖先と区別されるようにした。重要なのは、40万年の遺伝的変化にもかかわらず、彼らは依然としてクマであり、他の種に変わっていないということである。
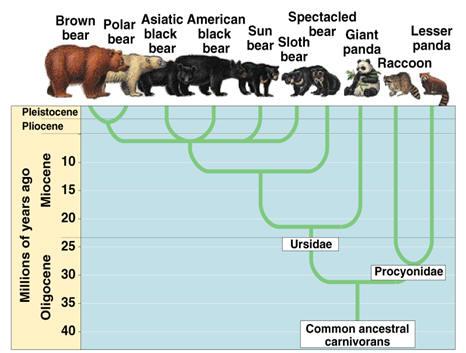
図3.10. クマの進化図
- フィンチのくちばしの遺伝的適応
ダーウィンのフィンチのくちばしの大きさと形の変化は、環境変化に対する遺伝的適応の古典的な例である。ガラパゴス諸島では、さまざまな食料源を利用するために、フィンチはくちばしの形を変化させてきた。干ばつが発生し、硬い種子が主な食料源となる場合、より大きくて強いくちばしを持つフィンチは選択的な優位性を持つ。逆に、環境が柔らかい餌を好むように変化すると、より小さく、より機敏なくちばしを持つフィンチが選択的優位性を持つようになる。
このような適応は、ALX homeobox protein 1(ALX1)遺伝子のようにくちばしの形に影響を与える特定の遺伝子と、High-mobility group AT-hook 2(HMGA2)遺伝子のようにくちばしの大きさに影響を与える遺伝子の変化の結果である。環境の変化は、これらの遺伝的変異に作用し、様々な生態学的環境に適したくちばしの形状をもたらす。何世代にもわたって、このような遺伝的適応は、フィンチが利用可能な資源を効率的に利用することを可能にし、環境変化が遺伝的変化をどのように誘導するかを示す良い例である。フィンチは約200万年前からガラパゴス諸島に生息している。この長い期間にもかかわらず、彼らはまだフィンチのままであり、他の種に変わるような大進化は起きていない。
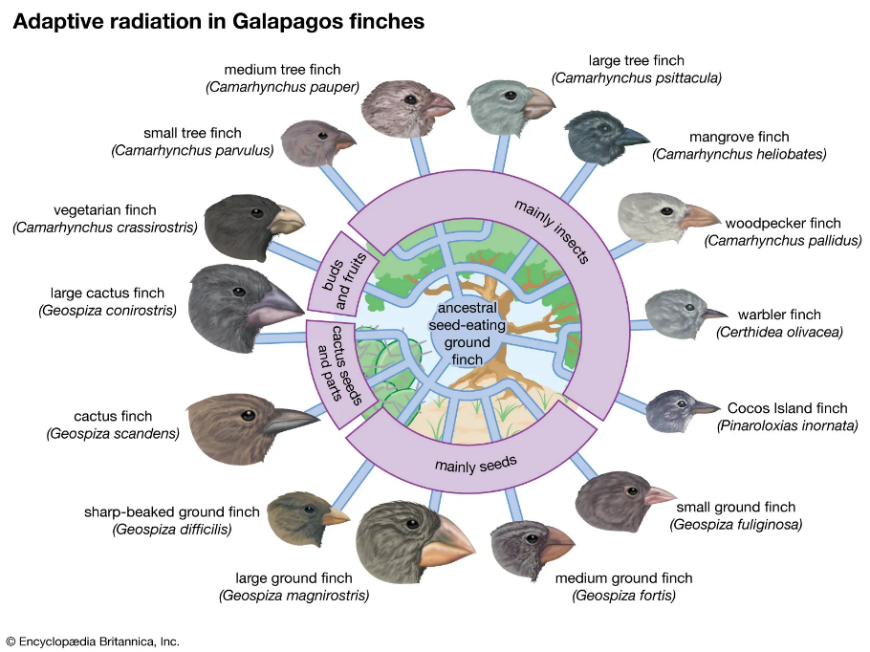
図3.11.変化する環境に遺伝的に適応したフィンチのくちばし
ダーウィンは《種の起源》で、自分の理論を「進化論」と明示的に主張していない。代わりに、彼は「自然選択による種の起源」をテーマに「変異」、「適応」、「自然選択」などの用語を使用した。「進化」という言葉は、《種の起源》の初版が出版された後、同時代の人々によって広く使われるようになり、以降の版で明示的に使われるようになった。いずれにせよ、この「進化」という用語は、上記のいくつかの例で示したように「適応」という言葉の方が適切であり、ダーウィンの理論は「進化論」と呼ばれるよりは「遺伝的適応論」と呼ばれるべきである。
d. 私たちは類人猿から進化したのか?
人類学者によると、人間の進化は約2千万年前にHominoideaから始まったと言われている。Hominoideaは HominidaeとHylobatidae(テナガザル)に分化し、HominidaeはHomininaeとPonginae(オランウータン)に、HomininaeはHomininiとGorillini(ゴリラ)に、HomininiはHomininaとPanina(チンパンジー)に、HomininaはAustralopithecus(アウストラロピテクス)とArdipithecusに分化した。人間は約250万年前にアウストラロピテクスからHomo habilis、Homo erectus、Homo sapienceを経て進化したと言われている。果たしてこれは本当なのだろうか?

図3.12.人間は類人猿から進化したのか?
前章で見てきたように、フィンチの場合、200万年経ってもまだフィンチのままであり、変わったのはくちばしの形だけである。しかし、ほぼ同じ期間である250万年の間にチンパンジーのような類人猿が多くの身体的変化を経験し、知能面でも飛躍的な発展を遂げて人間に進化したというのは、理解しがたい主張である。
この問題を詳しく調べるために、アウストラロピテクスが人間になるためにはどの程度の遺伝子変異が必要なのか、遺伝学的にアプローチしてみよう。
そのためにはアウストラロピテクスと人間の遺伝子地図が必要である。人間の遺伝子地図はあるが、アウストラロピテクスの遺伝子地図は持っていない。アウストラロピテクスの中で最もよく知られているルーシーの脳は、現代のチンパンジーの脳と似た大きさを持っている。したがって、アウストラロピテクスの遺伝子が人間に最も近いチンパンジーの遺伝子に似ていると仮定しよう。チンパンジーの遺伝子分析は2005年に初めて行われたが、人間の遺伝子と比較してみると、約1.2%の違いが見つかった。
親から次の世代に遺伝子が受け継がれる際、約60個程度の遺伝暗号にエラーが発生する突然変異が起こる。この数字は、1億個のDNA文字のうち、わずか1文字だけが変異する確率である。一世代を25年とし、アウストラロピテクスから人間まで進化するのに250万年がかかるとすると、この間に10万世代が経過し、遺伝子変異は10万倍1億分の1の0.1%の遺伝子変化が起こることができる。この0.1%(10万/100億)の遺伝子差は現生人類の様々な人種間の平均遺伝子差と似ており、チンパンジーレベルの類人猿が人間に進化するのに必要な遺伝子変異の約10%しかない。したがって、250万年の間にアウストラロピテクスから人間に進化したことは遺伝学的に不可能であることが分かる。この計算は、遺伝子変異が次の世代に有益な変異が現れることを前提に達した数値であり、少しでも有害な変異が一緒に現れたら、人間に進化することはさらに不可能な数値に達するだろう。
この問題は、遺伝子のランダムな突然変異によるコドン変異を考慮して考えることもできる。人間とチンパンジーは共に約20,000~25,000個のタンパク質をコードする遺伝子を持っている。各遺伝子は、代替スプライシングと翻訳後修飾により複数のタンパク質変異体を生成することができ、その結果、約8万~10万個のユニークな機能性タンパク質を生成することができる。人間のタンパク質のアミノ酸数は約20~33,000個になる。人間とチンパンジーの間に1%の遺伝子差があるとし、両種とも1つのタンパク質当たり平均100個のアミノ酸を持つ2万個のタンパク質コード遺伝子を持っていると仮定すると、チンパンジーが人間に進化するためには、チンパンジーの各タンパク質当たり1つのアミノ酸が人間のアミノ酸と一致するように変異しなければならない。
チンパンジーのDNAでこのような突然変異が発生するためには、64個の可能なコドンのうちどのコドンに変異してもよいが、stopコドン(UAA, UAG, UGA)への変異は避けなければならない。なぜなら、アミノ酸配列にstopコドンが入ると、ナンセンス変異(nonsense mutation)となり、タンパク質合成を止めて全く機能しない非機能性タンパク質になるからである。2万個のタンパク質でstopコドンとチンパンジー自身のコドンで突然変異が起こらず、人間のコドンに対応する1%の突然変異率を達成する確率は(60/64)20000 = 10-561 ≈ 0である。フレームシフト変異(ヌクレオチドの挿入または削除)を考慮しないとしても、この確率は非常に低く、ランダムな偶然によって発生することは事実上不可能である。この結果もアウストラロピテクスから人間に進化するような大進化はランダムな突然変異によって不可能であることが分かる。したがって、人間は猿から進化しなかったという事ができる。
e. 知的設計論
知的設計論(Intelligent Design)は、生命体を含む宇宙万物が偶然の産物ではなく、知的設計者である創造主神によって設計されたという理論で、創造論の婉曲的な表現である。このため、多くの進化論者たちが知的設計論を攻撃し、最終的には疑似科学に売り込んでしまった。このような結果、創造論者であっても知的設計論を受け入れない人が多い。しかし、知的設計は生命体と万物の創造に対する科学的な説明として、創造論の別の名前として受け入れなければならないと思う。
知的設計に関連する注目すべき事件は、2005年に米国ペンシルバニア州ドーバーで行われた連邦裁判である。この裁判は、保護者たちが公立学校で知的設計を教えることが憲法に違反すると主張して訴訟を起こしたことから始まった。保護者たちは、知的設計は本質的に宗教的な性格を帯びており、公立学校でこれを教えることは、政教分離を義務付けた米国憲法の設立条項に違反すると主張した。裁判が行われている間、知的設計と進化論を支持する人々はそれぞれの主張を展開した。知的設計を代表する生化学者マイケル・ビヒは、生命体の複雑な構造は自然選択だけでは説明できず、知的な原因によって形成されたことを提示した。しかし、裁判所はビヒと他の知的設計支持者の主張を拒否し、代わりに進化論支持者の主張を受け入れた。 そして、裁判官はドーバー公立学校で知的設計を教えることは違憲であると判決した。
この判決の最大の問題は、裁判所が進化論支持者の主張と関連する科学論文を無批判に受け入れたことにある。これらの論文は暗黙的に生命体がランダムな過程で偶然発生したと仮定し、生命体の環境に対する遺伝的適応を進化の証拠として誤って解釈したものである。しかし、表3.3に要約されているように、進化論は現存する生命体にのみ適用され、生命の起源を説明することはできない。また、進化論は生命体の遺伝子コードにすでに内在している環境変化に対する遺伝子の発現を単純に説明するだけで、その根本的な原因は説明できない。しかし、裁判所はこのような事実を考慮せずに判決を下した。
知的設計論の始まりは、1802年に出版されたウィリアム・ペーリーの著書《自然神学:自然の姿から収集した神の属性と存在の証拠》にある時計職人論証に遡ることができる。ペリーは、時計の複雑さが設計者を暗示するように、生命と宇宙の複雑さも神の創造者を暗示すると主張した。知的設計の主な概念には、特定複雑性、還元不可能な複雑性、微調整が含まれる。微調整の様々な例は前章で述べたので、この章では特定複雑性と還元不可能な複雑性について説明する。
i. 特定複雑性
知的設計の中心的な概念である特定複雑性は、自然界に見られる特定のパターンが非常に複雑でありながら、特定の機能を実行するために精巧に配置されているという考えである。ランダムな複雑性とは異なり、特定複雑性は単に複雑であるだけでなく、特定の結果を達成するために整然と配置されている。このような二重の性質は、そのようなパターンが単なる偶然によって発生したのではなく、目的を持って発生したことを意味する。例えば、1,000個のアルファベットを無作為に組み合わせれば、複雑であるが、意味は明確ではない。しかし、1,000個のアルファベットで文章を作ると、複雑な組み合わせのように見えるが、その意味は明確であり、何らかの意図を持ってその文章を作成したことが分かるのと同じ理屈である。
特定複雑性の代表的な例は、DNAの構造である。DNAのヌクレオチド配列は非常に複雑で、一本鎖でも無数の組み合わせを持つ。DNAの複製と修復メカニズムは、その複雑さをさらに強調する。このようなプロセスは、遺伝情報を正確にコピーして維持するために、複数のタンパク質と酵素が協力して動作する。ヌクレオチド配列は単に複雑であるだけでなく、タンパク質合成のための正確な指示を暗号化するほど非常に具体的である。DNA配列の各遺伝子は特定のタンパク質に対応し、配列のわずかな変化も、結果として得られるタンパク質の機能に大きな影響を与える可能性がある。また、DNAは遺伝子がいつ、どこで発現されるかを制御する調節要素を含んでおり、別の特異性を加えている。これらの事実は、DNAで観察される特定複雑性が、ランダムな突然変異や自然選択のような非指向的なプロセスによって生じた可能性は低く、代わりに知的設計がこのような洗練された機能的に特異な情報の起源をよりよく説明していることを意味する。
特定複雑性の別の例は、細菌の鞭毛である。細菌の鞭毛は、特定の細菌が運動に使用する鞭のようなモーター構造であり、フィラメント、フック、基底体などのさまざまなコンポーネントを形成する約40種類のタンパク質で構成されている。基底体自体は、ローター、ステーター、ドライブシャフト、プロペラを備えた回転エンジンのように機能する。
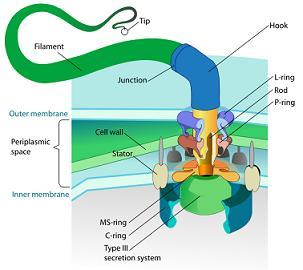
図3.13. 細菌鞭毛 の構成
鞭毛が機能するためには、これらすべての部品が存在し、正確に組み立てられている必要があり、これらのコンポーネントのいずれかが欠けても鞭毛は機能しない。鞭毛を動作するために非常に特殊なタンパク質は所定の順序で組み立てられ、その形状は、よく設計された機械の部品のように正確にフィットしなければならない。鞭毛は複雑であるだけではなく、細菌を推進する非常に特殊な機能を果たしている。驚くべき速度で動作し、方向を変え、エネルギー効率が高く、目的を持って設計されていることを示している。細菌の鞭毛の特定複雑性は、ランダムな突然変異や自然選択では適切に説明できず、このような高度に統合された機能的なシステムが偶然に発生する可能性は極めて低い。さらに、鞭毛の中間形態は非機能的である可能性が高いため、進化論が示唆するような緩やかな段階的進化では説明できない。
ii.還元不可能な複雑性
還元不可能な複雑性とは、生化学者マイケル・ビヒが提唱した概念であり、特定の生物学的システムは、段階的な段階的修正では進化できないほど複雑であるという理論である。このようなシステムには、視覚サイクル、細菌の鞭毛、血液凝固連鎖反応、細胞内輸送システム、抗原抗体反応、そしてAMP(アデノシン一リン酸)の生合成などがあり、基本的な機能をする多くの構成要素が相互作用しながら調和して構成されるシステムである。そして、その構成要素のいずれかが除去されると、そのシステムの機能がすべて停止することになる。このような精巧で相互依存的な構造は、自然選択とランダムな突然変異だけでは説明できず、知的設計者の設計によってのみ可能であることを示唆している。
還元不可能な複雑性の一例である視覚サイクル(visual cycle)について詳しく見てみよう。目の視覚サイクルは、光を電気信号に変換して視覚を可能にする生化学的プロセスである。視覚サイクルの主な構成要素は、光受容体(視間体と円錐体)、ロドプシン、オプシン、レチナール、信号伝達経路、神経処理プロセスなどである。光受容体は光を感知する網膜の細胞であり、間上体は弱い光を感知し、円錐体は色を感知する。各光受容体はロドプシンという光色素分子を含み、これはタンパク質であるオプシンと光に敏感な分子であるレチナールで構成されている。円錐体は、さまざまな波長の光に反応するさまざまなオプシンを含み、色覚を可能にする。レチナールはビタミンAの誘導体であり、光を吸収すると形状が変化する。この形状の変化はオプシンを活性化し、視覚変換の連鎖反応を開始する。活性化されたオプシンはトランスデューシンというGタンパク質を活性化し、トランスデューシンはPDE(ホスホジエステラーゼ)を活性化し、細胞内のcGMP(環状グアノシン一リン酸)レベルを下げる。cGMPレベルが低下すると、光受容体細胞膜のイオンチャネルが閉じられ、これは細胞の過分極をもたらし、電気信号を生成する。電気信号は正極細胞を介して神経節細胞に伝達され、視神経を介して脳に伝達され、脳はこれらの信号を処理して視覚イメージを形成する。このような一連の視覚サイクルは瞬間的な速度で行われ、非常に少ない光によっても効率的に行われ、暗いところでも物体をよく分別できるようにする。理解を助けるために、これらの過程のアルゴリズムをパイソンプログラムで表すと次のようになる。
コンピュータプログラムで表現された視覚サイクル
# 初期化: 視覚サイクルのための光受容体環境設定
class VisualCycle:
def __init__(self):
self.photoreceptors = {'rods':[], 'cones':[]}。
self.initialize_photopigments()
self.signal_pathway_active = False
# ユーザー入力: 入ってくる光を検出し、光色素活性化プロセスを開始する。
def detect_light(self, light_wavelength):
if light_wavelength in visible_spectrum:
self.activate_photopigment(light_wavelength)
#イベントの生成:レチナールの形状を変化させ、オプシンを活性化し、シグナル伝達経路を誘発する。
def activate_photopigment(self, wavelength):
retinal = self.change_retinal_shape(wavelength)
opsin = self.bind_retinal_to_opsin(retinal)
self.start_signal_transduction(opsin)
#イベント処理:トランスデューシンとPDEを活性化し、cGMPレベルを低下させ、イオンチャネルを閉じて電気信号を生成する。
def start_signal_transduction(self, opsin):
self.signal_pathway_active = True
transducin = self.activate_transducin(opsin)
pde = self.activate_pde(transducin)
self.regulate_cGMP_levels(pde)
self.generate_electrical_signal()
#信号処理: cGMPのレベルに応じてイオンチャネルを調整し、電気信号の生成を促進する。
def regulate_cGMP_levels(self, pde):
cGMP_level = self.reduce_cGMP(pde)
self.adjust_ion_channels(cGMP_level)
# 信号出力: 電気信号を生成し、脳に送信する。
def generate_electrical_signal(self):
if self.signal_pathway_active:
electrical_signal = self.create_signal()
self.transmit_signal_to_brain(electrical_signal)
#ネットワーク通信: 信号を処理し、バイポーラと神経節細胞を経由して伝達し、最終的に視神経を経由して信号を送信する。
def transmit _signal_to_brain(self, signal):
bipolar_cells = self.process_signal_with_bipolar_cells(signal)
ganglion_cells = self.forward_signal_to_ganglion(bipolar_cells)
optic_nerve = self.send_signal_via_optic_nerve(ganglion_cells)
self.visual_perception(optic_nerve)
#最終出力:脳が信号を解読して処理し、視覚イメージを生成する。
def visual_perception(self, optic_nerve):
visual_cortex = self.decode_signal(optic_nerve)
self.render_image(visual_cortex)
視覚サイクルをコンピュータプログラムで表した上記の例えから、我々は次のような事実を明らかにすることができる。視覚サイクルの各コンポーネントは相互に依存しており、光受容体、ロドプシン、レチナール、トランスデューシン、PDE、イオンチャネルが既に存在し、正常に機能している必要がある。もしいずれかの構成要素が存在しなかったり、視覚サイクルの順番が変わっても、視覚サイクルは機能しない。したがって、このような複雑なシステムは、すべての構成要素がない中間段階は機能しないので、小さな段階的な変化の連続による進化論では説明できない。視覚サイクルがコンピュータプログラムでうまく表現されることは、目が創造主によって知的に設計されたことを意味する。これが知的設計論の一例であり、目の設計図は創造主が設計された染色体11番にあるPAX6という目を作る遺伝子である。
それにもかかわらず、進化論者は目の起源を説明するために、十分な根拠のない仮説を提示している。ダーウィンは、無脊椎動物に見られる単純な視神経が段階的な変化を経て脊椎動物の複雑な目に進化したと主張した。ドーキンスもスウェーデンの生物学者の模擬実験を言及しながら、目は明暗と方向を感知する単純な組織から36万4000世代を経て水晶体を備えたカメラレベルに進化したと主張する。上記の実験は魚の目を対象としたもので、もし魚の一世代を10年とすると、視神経から完全な目に進化するのに約360万年かかるという話だ。この長い期間、見えない目でどのように摂食活動をし、捕食者からどのように生き残ったのか疑問である。そして、DNAやタンパク質さえも自然に形成することができないのに、最初の視神経はどのような過程を経て生成されたのかについての説明がない。また、目の進化が可能になるためには、視覚に関連する多数の遺伝子変異とタンパク質の相互作用がなければならない。しかし、単純な視神経から水晶体を備えた目に発展するまでに、このような遺伝子ネットワークがどのような過程を経て徐々に形成されたのか、分子生物学的証拠が不足している。例えば、レチナール、トランスデューシン、PDE、イオンチャネル、神経回路網などの要素が個別に進化したとしても、これらが調和して機能を発揮するには、同時に特定の方法で相互作用しなければならない。しかし、このような複雑な要素がランダムな過程を通じて適切に組み合わされて洗練された視覚機能を形成したという主張は、論理的に納得しにくい。
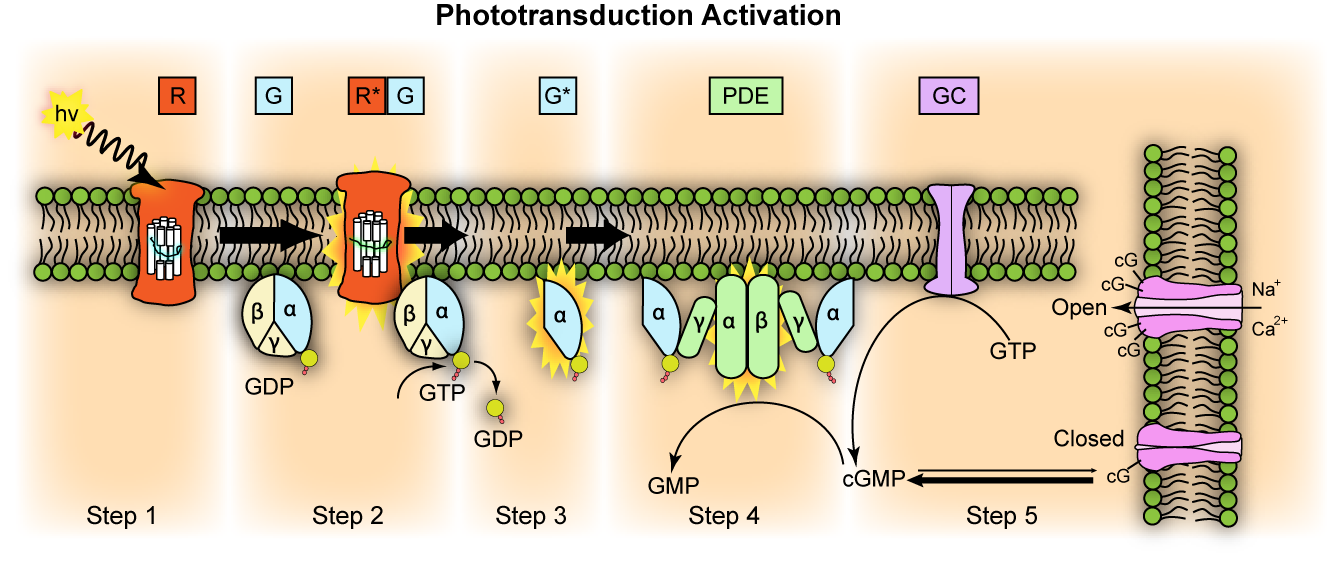
図3.14.ビジュアルサイクルの動作過程
iii. 知的設計論の主な書籍
この章では、知的設計論に関する重要な書籍を紹介する。
Evolution: A Theory in Crisis, Michael Denton, 1985: この本で、マイケル・デントンは、分子生物学と 古生物学など様々な分野の証拠を提示し、生命体で観察される精巧な構造と機能は、ランダムな突然変異と自然選択では説明しにくく、知的設計を指していると説明している。
Darwin's Black Box: The Biochemical Challenge to Evolution, Michael J. Behe, 2006: この本でマイケル・ビヒは還元不可能な複雑性の概念を紹介し、細菌の鞭毛のような特定の生物学的システムが自然選択や段階的進化では説明できないほど複雑であり、知的設計の産物でなければ説明できないと主張する。この本がアメリカで最初に出たとき、NatureやScienceのような科学的なジャーナルはもちろん、Wall Street Journalのような比較的一般的なジャーナルに至るまで、多くの場所で批判された。ビヒの本は、創造論を知的設計という科学の域に引き上げたと評価される。
Darwin on Trial, Phillip Johnson, 2010:フィリップ・ジョンソンによって書かれた審判台のダーウィンは、知的設計運動の出発点であり、ダーウィニズムに対する学術的な批判と新しい科学のパラダイムとして知的設計運動を開いたと評価されている。この本の第1章は法的背景としてこの本の執筆目的を述べており、第2章から第8章まで自然選択、大小の突然変異、化石問題、進化の事実、脊椎動物の系統、分子的証拠、生命以前の進化など進化論に関連する直接的な内容を扱っており、第9章から第12章までは科学のルール、ダーウィン主義の宗教、ダーウィン主義の教育、科学と疑似科学など進化論に関連する哲学的な内容を扱っている。
Signature in the Cell: DNA and the Evidence for Intelligent Design, Stephen C. Meyer, 2010: この本は、細胞に対する現代分子生物学と生化学の発見に基づき、地球上の最初の生命の出現は知的存在の設計によって可能であったことを論じている。彼は、DNA内の遺伝情報の精巧さに基づき、知的設計に関する詳細な事例を提示し、生命の起源がランダムなプロセスではなく、 目的のある創造を指していることを示唆している。
Darwin Devolves: The New Science About DNA That Challenges Evolution, Michael J. Behe, 2020:Beheの別の本で、最近の遺伝的発見が伝統的なダーウィンの進化論を弱体化させると主張している。彼は、自然選択とランダムな突然変異は小規模な適応を説明できるが、細胞内の分子機構を説明するには不十分であると主張している。ビヒは「退化」という概念を導入し、突然変異が新しい有益な特性を生み出すのではなく、遺伝情報を失う原因となり、これらの遺伝的限界が知的設計者の必要性を示していると主張し、知的設計が生命の複雑さを説明するのに適していることを示唆している。
The Mystery of Life's Origin: Reassessing Current Theories, Charles B. Thaxton et al., 2020: この本は、生命の起源を説明する様々な進化論的理論の欠点を議論し、知的設計を科学的に有効な代替案として紹介している。現代知的設計運動の基礎を築いた本と評価される。
The Design Inference: Eliminating Chance through Small Probabilities, William A. Dembski & Winston Ewert, 2023: この本は、自然界で知的設計を検出するための理論的基礎を提供する。特定複雑性、小さな確率、フィルタリング技術の3つの主要な概念を使用して、知的設計を推論している。ある現象が複雑であると同時に特定の機能を実行するとき、または自然現象がランダムに発生する確率が非常に低いときは、知的原因によって設計されたことを示す。フィルタリング技法は、特定の現象が自然法則によって説明できるかどうかを確認し、もし自然法則で説明できない場合、第二段階では、ランダムな過程によって説明できるかどうかを検討する。ランダムな過程でも説明されない場合、最後の段階で知的設計によって説明される可能性を考慮するのが フィルタリング技法である。複雑な生物学的システムは、ランダムなプロセスによって形成される可能性が低いため、知的原因によって設計されたことを示す。
f. 粒子物理学と創造論
先ほど、原子、アミノ酸、RNA、タンパク質、DNA、細胞などで構成される生命体の起源について見てきた。そして、この議論では、生命体を構成している最小単位である原子は当然存在することを前提に出発した。では、天地万物を構成している原子はどのようにして存在するようになったのだろうか?単純な偶然の結果なのか、それとも創造主によって創造されたのだろうか?ここでは、天地万物の根源である原子を構成している素粒子(小粒子)はどのように生成されたのか調べていこう。
粒子物理学の標準模型によると、この世界のすべての物質は、6つのクォーク、6つのレプトン、4つのゲージボソン(グルーオン、光子、Zボソン、Wボソン)、ヒッグスボソンなど17個の基本粒子で構成されている。各粒子は特定の質量、電荷、スピンを持っており、まるで細胞の中でそれぞれの役割を忠実に遂行する細胞小器官のように、粒子の相互作用において特定の役割を果たす。
クォークは物質を構成する重要な骨格で、2つのアップクォークと1つのダウンクォークが強力に結びつけば陽子になり、1つのアップクォークと2つのダウンクォークが強力に結びつけば中性子になる。強力は、クォークがグルーオンを授受しながら生成され、距離の逆二乗に比例する重力や電磁気力とは異なり、核子内では距離が離れると力が強くなり、距離が近づくと力が弱くなり、クォークが一定の距離を維持するようにする。クォークは粒子の相互作用の中で他のクォークに変わるが、例えば、中性子がベータ崩壊で陽子に変わると、中性子にあったダウンクォークがアップクォークに変わって陽子になる。このようにクォークが変わるパターンは数学的にCKM行列に従う。
図3.15.標準モデルの基本粒子たち
基本粒子はヒッグスメカニズムによって質量を付与されるが、任意の質量値を付与されるのではなく、各粒子ごとに必ず決まった質量値を付与される。粒子が質量を付与されるときにヒッグス粒子が生成されるが、この粒子の実体が2012年に確認され、この発見でヒッグスとアングレールがノーベル賞を受賞した。
ゲージボソンは粒子の相互作用中の力を媒介するが、光子は電磁力、WボソンとZボソンは弱い力、グルーオンは強い力を媒介する。この粒子間の相互作用は、量子力学、量子場論、量子色力学などを通じてすべて数学的に記述することができる。
粒子物理学の世界は広大で少し難解だが、上記の簡単な事実だけを見れば、次のような根本的な疑問を投げかけることができる。
- 万物を構成する17個の基本粒子はどのように作られたのか?エネルギーから偶然に抽出されたのだろうか?それとも精密な数学的設計によって生成されたのか?
- ゲージボソンはどのようにして力を媒介する特性を獲得したのだろうか?たまたまこのようなメカニズムが生成されたのだろうか?それとも精密な数学的設計の産物なのか?
- 基本粒子ごとに決まった質量を付与するヒッグスメカニズムは自然に生まれたのか、それとも精密な数学的設計の産物なのか?
- ベータ崩壊で中性子が陽子に変わる過程は自然なのか?それとも数学的設計の産物か?
- 粒子のすべての相互作用はどのようにして数学的に記述可能なのか?偶然なのか、それとも数学的原理に基づいて設計されたからなのか?
この5つの事実のどれか一つでもランダムなプロセスによる偶然の結果であれば、この世界は存在しなかったかもしれない。なぜなら、基本粒子の一つでも欠けていたり、ヒッグスメカニズムが確立されていなかったり、基本粒子の質量とスピンの値が少しでも違えば、電子、陽子、中性子などが作られなかっただろうし、たとえ作られたとしても、互いにしっかりと結合することができなかっただろう。もしそうであれば、世の中のすべての物質は崩壊し、人間を含むいかなるものも形成することができなかっただろう。ここで私たちは、粒子物理学の世界でも知的設計論の還元不可能な複雑性を観察することができる。
物質を形成するための基本粒子の生成は、生命体形成のための細胞と細胞小器官の生成に例えることができる。特定の細胞と細胞小器官がそれぞれ固有の役割と特性を持っており、生命体を作り、維持するように、基本粒子もそれぞれ固有の役割と特性を持っており、原子、分子、そして最終的には物質を形成し、維持する。このような類似点は、細胞の微視的レベル、物質の小粒子レベル、生命体、星、銀河などの巨視的レベルなど、すべての自然界に知的設計の概念が含まれていることが分かる。
基本粒子の存在と相互作用が複雑な量子力学の数式で表現できるということは、これらすべてが数学的に精巧に設計されたことを意味する。そうでなければ、それぞれの素粒子が考える能力があり、どのようにすれば物質を構成できるかを知り、正確な質量、電荷、スピンの値を持って生成され、お互いの相互作用で複雑な数学的原理を使って行動することを受け入れなければならない。この事実は、思考能力のない素粒子が難しい量子力学の原理を完璧に理解し、適用していることを受け入れなければならないが、これは事実ではないことを誰でも知っているからだ。
g. エイリアンと創造論
宇宙にある無数の星の中には、私たちの太陽系と似た条件を持つ星が数え切れないほどあり、そのような星の周りを回っている惑星のうち、私たちの地球と似た条件を持つ惑星も無数にあるはずだ。したがって、進化論が正しいなら、私たちの地球だけでなく、地球と似た条件を持つ無数の惑星でも生命体が発生したはずだ。その場合、ある惑星では生命体が発生したばかりで、バクテリアレベルの生命体が住んでいるだろうし、ある惑星ではずっと前に生命体が発生し、現在私たちが想像もできない高度な文明を享受しているだろう。もし私たちがこのように多くの宇宙生命体の存在を発見すれば、創造論と進化論の議論に終止符を打つことができるだろう。
銀河系内の地球外生命体が住める惑星の数Nは、次のドレイク方程式によって推定することができる:
N = R*× fp× ne× fl× fi× fc× L
ここで、R*は星の誕生率、fpは星が惑星を持つ確率、neは生命体が現れることができる惑星の数、flは実際に生命体が誕生する確率、fiは生命体が知的生命体に進化する確率、fcは宇宙に信号を送ることができる文明の割合、そしてLはその文明が持続する期間である。各変数に適切な値を適用すると、1つの銀河あたり外系 文明が存在する惑星は約2個と推定される。

図3.16.エイリアンは存在するか?
宇宙にエイリアンの存在の有無を直接的に調べるエイリアン探査計画(SETI計画)は、1960年代初めにドレイクによって始まった。初期に行われたエイリアン探査計画はオズマ計画とオズマ計画IIと呼ばれ、米国グリーンバンクにある電波望遠鏡を使用して地球に近い約650個の太陽型星を観測した。その後、NASAで行われたサイクロプス計画、その後に行われたフェニックス計画、バークレー大学で行われたSERENDIP、SETI@home、アレン電波望遠鏡計画、ハーバード大学とプリンストン大学で行われた光学SETI計画などを通じて、太陽に似た1,000個以上の星がさらに観測された。 では、その結果はどうなったのだろうか?

図3.17.SETIに使用された電波望遠鏡
これまで60年、様々なSETI計画を通じて宇宙人の存在を追跡してきたが、その存在は発見できなかった。最近、百万個の星と百個の銀河を観測し、人工知能で分析した'画期的な聴音(Breakthrough Listen)計画'の結果もやはり異星人文明を発見できなかった。では、エイリアンは存在しないのだろうか?
エイリアンの存在は確認されていないが、もし多数のエイリアン文明が存在するのであれば、彼らはすでに私たちを訪問したか、あるいは今訪問しているかもしれない。その場合、彼らはどのような方法で宇宙旅行をするのだろうか?私たちが一般的に考えているUFOのような飛行物体に乗って広大な宇宙を旅することはほとんど不可能だ。なぜなら、飛行機より30倍も速いロケットで飛んでも、最も近い星であるプロキシマ・センタウリまで行くのに15万年かかるからだ。このような広大な宇宙を短時間で旅行する方法を見つけなければならないが、可能な方法としてはワープドライブやワームホールを通じた旅行が考えられる。
図3.18.ワームホール
ワープドライブは、アインシュタインの一般相対性理論にインスパイアされ、光より速い宇宙旅行のための理論的概念である。1994年に物理学者ミゲル・アルクビエレが提案したワープドライブは、宇宙船の前の空間を収縮させ、後ろの空間を拡張する「ワープバブル」を生成し、相対性理論に違反することなく、外部観測者と比較して光よりも速く移動できる方法だ。理論的には可能な方法だが、実際に使用するためには負のエネルギー密度を持つ物質が必要なため、現時点では実現可能性は低い。
アインシュタインの一般相対性理論によって予測されたワームホールまたはアインシュタイン-ローゼン橋は、広大な宇宙の距離をワームホールを通して瞬間的に旅行することができる。しかし、実際にこの方法を使用するには、通過するワームホールを安定化する必要があり、ワームホールの崩壊を防ぐためにワープドライバーのように負のエネルギー密度を持つ物質が必要である。
宇宙を瞬間的に移動するためのもう一つの方法は、テレポーテーションと呼ばれる瞬間移動または空間移動である。ブレーン宇宙論(Brane cosmology)によると、私たちの3次元宇宙はより高い次元である超空間(bulkまたはhyperspace)の一部であるということである。超空間を通じた瞬間移動は、既存の3次元空間を迂回して広大な距離を瞬間的に移動できる方法である。

図 3.19 . Teleportation
最近の観測によると、最初の銀河と星は今から約134億年前に形成されたが、進化論が正しい場合、すでにその時から生命体の進化が始まっているはずだ。したがって、ある生命体は私たちより1億、10億年、あるいは最高94億年(私たちは40億年前から進化を開始した)先に進化を開始し、今は私たちが想像できないほどの高度な文明を形成しているはずだ。
このような生命体が134億年前から宇宙の様々な場所で均一な頻度で進化を開始したと仮定すると、約70%(94億年/134億年)程度の宇宙生命体が私たちより先に進化を開始し、そのうち90%は私たちより9億4千万年先に進化し、99%は私たちより9千4百万年も早く進化を開始したことになる。今、極端に保守的な立場を取って、彼らのうち99%ではなく1%だけ、また、私たちより9千4百万年ではなく、わずか100万年先に進化したと仮定してみよう。
その場合、先のドレイク方程式で計算された千億個の知的生命体のうち1%である40億個の知的生命体が、私たちより100万年先に進化を開始したことになる。彼らが私たちより100万年前に私たちと同じような進化経路に沿って進化したと仮定すれば、彼らの文明は私たちの文明より100万年以上進んでいるはずだ。このように100万先に進化した異星人文明体は、宇宙旅行のための瞬間移動方法をすでに発見しているだろうし、今頃は私たちが隣国を訪問するように、宇宙のどこへでも瞬間的に旅行することができるだろう。
そのような文明体の1つの文明体に10億人のエイリアンが住んでいるとすると、地球を隣の家のように瞬時に訪問できる全体のエイリアンの数は40億個の知的生命体 ×10億人で400京なることが分かる。そのうちの全員が一ずつ地球を訪問するなら400京人、1%だけ訪問すれば4,000兆人の宇宙人が私たちの地球を訪問することができるだろう。もし彼らが1日に1回地球を訪問するなら4,000兆人の宇宙人が、1年に1回なら11兆人、10年に1回なら1兆1,000億人の宇宙人が毎日地球を訪問することになるだろう。そうなれば、私たちの地球は、地球に住む人間よりもはるかに多くのエイリアンが押し寄せてきて、まさに足の踏み場もないほどの長蛇の列をなすことになるだろう。しかし、過去にも、そして現在もそのようなエイリアンの訪問の痕跡が全くないことはどう理解すべきなのだろうか?
エイリアンの地球訪問の痕跡がないことについて、1950年にエンリコ・フェルミが疑問を持ち、「みんなどこにいるのか?」と尋ねたフェルミパラドックス(Fermi paradox)として知られている。これに対する説明としては、i)エイリアンはすでに私たちのそばに来ているが、私たちが知らないだけ、とii)エイリアンは存在せず、宇宙には私たちだけである、という。i)の場合、世界の様々な観光地で地球人よりはるかに多くのエイリアンを見ることができるだろうし、ii)の場合、エイリアンの数を推定したドレイク方程式で、進化論に基づく「惑星で生命体が自然に生まれる確率」がゼロになるだろう。エイリアンがバクテリアや透明人間でなければ、何らかの形で私たちに彼らの存在が明らかになるはずなのに、まだ彼らの存在を確認できていないという事実は、ドレイク方程式で仮定した進化論が正しくないことを示唆している。
h. 生命体と本能と創造論
コンピュータはハードウェアとソフトウェアだけで構成されているのではなく、その中間形態であるファームウェアも含む。ファームウェアは、コンピュータの電源を入れるときに起動するBIOSまたはUEFIプログラムで、コンピュータのハードウェアおよび入出力装置を制御するプログラムが含まれている。つまり、コンピュータを作った人がそのコンピュータがどのように動作すべきかを指示する事項が一つ一つ記録されているもので、生命体が自然界でどのように生存すべきかが記録されている生命体の本能と同じ役割を果たすと見ることができる。コンピュータ設計者がROMにファームウェアを内蔵したように、本能は創造主が生命体の脳と神経系に内蔵したもので、生命体がランダムな結果として作られたのではなく、コンピュータのように創造主によって創造されたことを示唆する。それでは、生命体に内在するこの本能が生存のためにどのように実行されるのか、マメコバチとハタオリドリ(Weaver bird)の巣作りを通して考察することにしよう。
i.マメコバチの巣作り
マメコバチの巣作りはファーブルの昆虫記に詳しく書かれているが、内容をまとめると次のようになる。マメコバチは川辺の砂浜の平らな石を選び、その上に唾液で練った土塊を運んでドーナツ状に積み上げ、丈夫な巣を作る。その後、建てた家に蜜と花粉を入れ、卵を一つ産んだ後、巣の入り口を塞いでしまう。ここからファーブルは面白い実験を行う。
マメコバチが巣作りの最初の段階である土のドーナツを積んで作った後、再び土塊を取りに行ったとき、ほぼ完成した他のマメコバチの巣と入れ替えたら、土塊を噛んで戻ってきたマメコバチはそれを見てどのように行動するのだろうか。その場合、土塊を捨ててほぼ完成した巣に蜂蜜を詰めるのが正常な行動だが、そうせず、完成した巣の上に持ってきた生地を塗って巣作りを続ける。
図3.20. 完成した家に泥を塗って巣作りを続けるマメコバチ
これとは逆に、巣が完全に建てられ、蜂蜜が貯蔵されている巣を、蜂がいないときに基礎工事だけの巣と交換しておいた場合、蜂蜜を集めて巣に戻ってきた蜂はどのように行動するのだろうか。持ってきた蜜を下ろして、代わりに土の塊を持ってきて巣を完成させるのではなく、すぐにあふれそうなほど低い巣に、蜜を貯め続ける。

図3.21.基礎工事を終えた家に蜜を貯め続けるマメコバチ
この二つの実験を見ると、マメコバチは変化する状況と関係なく、本能に内在するプログラム通りに巣作りを行い、蜜を集める順番 に入り、その順番は変えることができないことが分かる。
ファーブルが行ったもう一つの実験は、マメコバチが蜜を蜂の巣に入れる順序と、卵から目覚めたマメコバチの行動である。マメコバチが蜜を持って巣に着くと、まず蜜袋の中の蜜を吐き出すために頭を巣箱の中に入れ、蜜を吐き出し、頭を抜いた後、代わりに尻を入れ、体についた花粉を払い落とす。しかし、蜂が蜂の巣に蜜を吐いて尻を入れる瞬間、尻を入れられない巣の外に追い出したら、どのような行動をとるのだろうか。その場合、蜂は再び飛んできて、少し前に蜜を吐いて体の中の蜜袋が空になっているにもかかわらず、再び頭から先に入れ、蜜を吐いてから尻を入れるという動作を先行する。
この実験も、マメコバチが本能に内在する蜜を満たすプログラム通りに行動し、その順番は変えられないことを示している。
図3.22.マメコバチは先に蜜を吐き出し、花粉を飛ばす(左)。花粉を払い落とすマメコバチを邪魔すると、再び蜜を吐き出し、花粉を払い落とそうとする(右)。
マメコバチが卵から目覚め、密閉された巣を突き破る行動でも同様の本能を観察することができる。マメコバチは巣作りが終わると蜜を詰めてその上に卵を産み、巣の入り口を塞いでしまう。ブロックされた入り口はセメントのように硬いのだが、卵から目覚めた成虫はすぐに大きくて丈夫な顎でその硬い壁を突き破って外に出て蜜を探しに飛んでいく。しかし、蜂の巣の入り口に円錐形の紙の蓋をかぶせると、硬い壁を突き破った成虫はどのように行動するのだろうか?セメントのように硬い壁を突き破ることができる顎を持って出てきたが、それよりも突き破りやすい紙の蓋にぶつかったら、紙の蓋を突き破ろうとする努力を全くせず、そのまま餓死してしまう。

図3.23.家にくっついた紙は突き抜けるが、円錐紙はどうしたらいいのかわからず餓死する。
ここから分かるのは、マメコバチは生まれてすぐ、硬い巣の入り口を一度だけ突き抜ければ、いくらでも花を探しに飛べる空間が広がるという期待が本能に内在していることが分かる。しかし、不意に紙で塞がれた小さな空間に出くわすと、その後はどうしたらいいのかわからず、そのまま餓死してしまう。この事実も、マメコバチが生まれた後に取るべき行動が、歯車のように噛み合っており、本能による順番通りに進まなければ自然界で生き残れないことを物語っている。
ii.ハタオリドリの巣作り
アフリカや西南アジアで見られるイエローハタオリドリの巣作りを見ると、マメコバチが見せたのと同じような本能を見つけることができる。ハタオリドリはちょっと変わった巣箱を作るのだが、草を一本一本噛んで編んでひょうたんのような形の巣を作る。

図3.24.ハタオリドリの巣
Maraisは、ハタオリドリのこのような複雑な巣作りの技術が純粋に本能的なものなのか、それとも学習された行動と関係があるのかを調べるために、興味深い実験を行った。生まれたばかりのハタオリドリをケージに閉じ込めて飼育し、産卵期に家を作らないようにし、卵を産むようにした。そうして生まれた子にも家を作るための草を与えず、床に卵を産ませるという実験を4世代目まで同じように繰り返した。その後、第5世代に生まれた鳥が成長し、産卵期になると、再び家を建てられる草を提供したところ、その鳥は本能的に写真の巣のような家を作り始めた。一度も見たなく、作り方も分からない家を設計図を見るように、どうしてこんなに完璧に作れるのだろうか?
ここで私たちは、コンピュータを作り、そのコンピュータがどのように動作するかを記録したファームウェアと、ハタオリドリを創造し、自然界でどのように生きるかを本能の中に記録した創造主の設計が同じ概念であることが分かる。
iii.オウムガイの巣作り
オウムガイ(Nautilus)は、熱帯地方の海で見られるサイズの貝である。オウムガイの貝殻は完璧な対数螺旋(Logarithmic spiral)に沿った美しい殻を作りながら生きている。どうしたことだろうか?誰もオウムガイが高校レベルの数学を学んで家をそのように作ったとは思えないだろう。

図3.25. 対数螺旋模様のオウムガイの殻
オウムガイがどのようにしてこのような素敵な貝殻を作るのか見てみよう。まず、オウムガイはマントルと呼ばれるところからカルシウムが豊富な液体を分泌して、前にあった殻より少し大きい直径の円形の殻を作り、貼り付ける。しかし、単純に直径が少し大きい円形を貼り付けると、貝殻が一直線に進み、ラッパのような形になり、丸く丸まった貝殻の形にはならない。ここでオウムガイは驚くべき技術を発揮して、円形の貝殻を作り、貼り付ける時、中心に近い円の下部よりも円の上部に分泌物を少し多く分泌する。そうすると、回転効果をもたらし、積み重なる円が直進せず、対数螺旋形の貝殻が作られる。
ここで私たちは次のような疑問を持つ。オウムガイはカルシウムが豊富な液体を分泌して家を建てるのに、どのようにしてそのような液体が必要なのか?分泌する液体の濃度、分泌速度、液体から硬い殻に硬化させる方法など、そのような精巧な生化学的過程をどうやって知ったのだろうか?オウムガイの意志なのか、それとも本能の中に記録された順序によって遺伝子発現が調節され、ある細胞ではカルシウムの液体が分泌され、別の細胞では分泌された液体が硬化するタンパク質が生成されるなど、様々な過程が調節されるのだろうか?
これに答えるために、3-Dプリンター(=オウムガイ)でオウムガイの貝殻を作るとしよう。3-Dプリンターを初めて購入したら、input無しで機械自体は何もしない。オウムガイである3-Dプリンターを作動させるためには、3-Dプリンターに使われるGプログラミング言語で、どのような順序と過程を経て貝殻を作るかを指示するプログラムが必要である。また、オウムガイの貝殻が作られる物質を液体の状態でインクカートリッジに注入しなければならない。その後、完成したプログラムを入力して3-Dプリンターを駆動すると、プリンターは入力されたプログラムが指示する通りにインクの噴射速度を調整して少しずつ貝殻を作っていく。
この単純な例えから見られるように、3-Dプリンターであるオウムガイは自分の意志で殻を作ることができず、本能の中に入力されたGプログラミング言語のような巣作りプログラムが指示する通りに殻を作る。この事実は、オウムガイの殻が偶然による進化の産物ではなく、創造主がオウムガイの本能と遺伝子に組み込んだ巣作りプログラムの産物であることを示している。
i. 自然に発見される数学的原理と創造論
「数学は神が宇宙を書き記した言語です。」 Galileo Galilei
私たちが自然界や動植物の成長形態をよく見ると、多くの部分で数学的な原理を発見することができる。自然界に現れた数学的原理の中で代表的なものは、黄金比、黄金角、フィボナッチ数列、対数螺旋、フラクタルなどである。まず、これらの数学的原理を簡単に説明しよう。
- 黄金比は、ある2つの数の比率がその和と2つの数のうち大きい方の比率に等しくなる比率(a/b=(a+b)/a)で無理数であり、近似値は1.618程度になる。
- 黄金角: 円の長い円弧と短い円弧が黄金比を成すときに形成される角度で、無理数であり、近似値は137.51度程度になる。
- フィボナッチ数列 : 0または1から始まり、その後ろのすべての項は直前の2項の合計である数列(例えば、0, 1, 1, 1, 2, 3, 5, 8, ...)。
- 対数螺旋: 極座標で半径が角度によって指数的に増加し、中心から引いた接線が常に一定の角度をなす螺旋で、フィボナッチ螺旋と似た形状である。
- フラクタル: 部分が全体と似た形を繰り返し持ち、拡大しても同じ構造が現れる自己類似性(self-similarity)を持つ幾何学的構造を指す。
図3.26.黄金比、黄金角、対数螺旋、フラクタル
それでは、これらの数学的原理が自然どこで見られるのか見てみよう。
葉序学(Phyllotaxis)は、植物の茎から葉が出る葉序を研究する学問である。葉書学によると、葉はネジの鋸歯のように螺旋状に回って出てくるが、葉がどれだけ回って最初に出た葉と同じ方向から出てくるかを示すことを開度(divergence angle)という。例えば、葉が1-2-3-4-5の順に葉が出ながら2周回って6番目に1番目と同じ方向から再び葉が出る場合、2周である720度を5回で回るので開度は720/5=144度であり、葉が1-2-3-4-5-6-7-8の順に葉が出ながら3周回って9番目に1番目の葉の方向から再び葉が始まる場合、葉は3周である1,080度を8回で回るので開度は1,080/8=135度である。2つの例のように、葉が2周を5回で回れば2/5葉序と呼び、3周を8回で回れば3/8葉序と呼ぶ。ところで、植物の葉は何の順番で生えるのではなく、1/2、1/3、2/5、3/8、5/13...の順番で生えるが、これをよく見ると、分母と分子がすべてフィボナッチ数になっていることが分かる。また、分母は分子のフィボナッチ数の次の次のフィボナッチ数になる。葉序の数が多くなると、開度は黄金角である137.51度に近づく。開度が黄金角に近づくと、葉序の重複が最小になる。このように葉序の重複が最小になると、どのような葉でも日光をまんべんなく受けることができ、雨や露が葉によく集まり、茎に乗って根に降りてきて、その植物の生存に不可欠な水をよく供給できる特徴を持つようになる。
図3.27.2/5葉序(a)と3/8葉序(b)
木の枝や草の葉が成長する順番もよく見るとフィボナッチ数列に従うが、その代表的な例がノコギリソウである。下の図にあるノコギリソウは最初に1本の枝が出た後、2本になり、その後3本、5本、8本、13本の順に枝が生え、各枝についた葉の数もフィボナッチ数列に沿って1本、1本、2本、3本、5本、8本の順で伸びる。
図3.28.ノコギリソウの枝と葉
植物の葉だけでなく、芽、実、種子もフィボナッチ数列と黄金角の支配を受ける。
ノルウェートウヒの芽吹きパターンは、フィボナッチ数列と黄金角の原理に従う。次の図に示すように、中心から角度を測ってみると、各芽は前の芽から約137.5度(黄金角)の角度で出てくる。その結果、枝が幹を中心にフィボナッチ数列の分布と一致するスパイラルパターンに形成される。この自然なパターンは、木が日光、水、栄養分を効率的に集めることができ、木を健康に成長させる。
図3.29.ノルウェートウヒの発芽パターン
ヒナギクの花びらと種子はフィボナッチ数列に沿って螺旋状に並んでおり、各方向の螺旋数は一般的に21、34などの連続したフィボナッチ数である。また、連続した花びらや種子間の開度はおおよその黄金角を保ち、その巻き模様は対数螺旋となる。対数螺旋は自己類似性という固有の特性があるため、デイジーの花びらが成長しても花びらの形がせず、美しい形をそのまま維持する。
松ぼっくり、カリフラワー、ロマネスコブロッコリーにも同様のパターンが見られる。松ぼっくりの鱗はフィボナッチ数に従って螺旋状に配置されており、一般的に一方向には8本、反対方向には13本の螺旋で巻かれており、各鱗はおおよその黄金角で配置されている。同様に、カリフラワーの花芽は一方向には5本の螺旋、他方向には8本の螺旋で巻かれており、ロマネスクブロッコリーの花芽は一方向には13本、他方向には21本の螺旋で巻かれている。
パイナップルは、目の配列からフィボナッチ数列を見つけることができる。パイナップルの目に沿って左上へ上がる螺旋と右上へ上がる螺旋、縦につながった線を数えてみると、列の数がそれぞれ5、8、13または8、13、21のようにフィボナッチ数列で連続した三項となる。このような配列は、効率的な空間活用と構造的安定性を最大化し、栄養分を均等に分配して均一な成長を助ける。

図3.30.植物に見られるフィボナッチ数列と対数螺旋
対数螺旋に沿った成長曲線は、植物だけでなく、人をはじめとする様々な動物にも見られるが、人の耳たぶ、耳の中にある蝸牛、人の指、海馬の尾、ヤギの角、そしてオウムガイをはじめとする様々な種類の貝やカタツムリの貝殻などだ。このように動植物の様々な器官や形が対数螺旋の形で成長しなければならない重要な理由があり、それは対数螺旋で成長する時だけ、幼い頃の姿と同じ姿を維持しながら成長し、その器官が持つ固有の機能が大きくなってもよく維持できるからである。
例えば、人の耳介は音をよく集め、耳の中の蝸牛は低音を十分に聞こえる強さに増幅してくれる最適な幾何学的形状を持っている。しかし、もしこのような重要な器官が成長するにつれてその形が変わるとしたらどうなるだろうか?その場合、子供の頃は耳がよく聞こえるが、大人になると耳が聞こえなくなるという困った状況にぶつかることになる。また、ログ螺旋に沿って成長する指の節が、子供の頃は拳を自由に握ることができるほど適当な比率だったが、成長するにつれてログ螺旋の形から外れると、拳すら握りにくい形の手をことになるだろう。

図3.31.蝸牛、耳、海馬、橈骨、指節骨
自然界で発見されるもう一つの数学的原理であるフラクタル構造は、シダの葉、木の枝、ロマネスクブロッコリー、そしてカウリーフラワーなどに見られる。私たちがシダの葉をよく見ると、葉の全体的な形がそれより小さい一つ一つの葉にコピーされて現れており、またそれよりさらに小さい葉でも全体の形が繰り返される構造を見ることができる。

図3.32.フラクタル構造を持つロマネスコブロッコリー
フラクタル構造は植物だけでなく、私たちの体の循環系、神経系、消化器官などにも見られる。循環系は新鮮な酸素と栄養分を体のすべての細胞に迅速に供給する役割を果たさなければならない。そのためには、体内の多数の細胞まで血管が分布しなければならないが、もしこれを通常の方法で設計すれば、私たちの体を血管だけで完全に満たさなければならないだろう。ところが、私たちの血管がフラクタル構造を持つようにすれば、最小限の体積を占めながらも最大の面積を持つことになる。また、血管がフラクタル構造を持つ場合、心臓の強力な拍動を大小多数の血管に分散させながら効率的に緩衝する役割を果たすことができる。
人間の心肺器官もフラクタル構造になっているが、肺の大きな枝は小さな枝に分かれ、小さな枝はそれよりさらに小さな枝に分かれ、最後に肺胞に至る。このようにフラクタル構造で形成された肺の表面積はコートほどの大きさになり、肺に入った空気から最大限の酸素を吸収させる。消化管の内部も栄養分を効率的に吸収するために微細なしわを持つフラクタル構造になっているが、その表面積はテニスコートの4倍程度になる。
自然や生物学的システムで見られる黄金角、対数螺旋、フィボナッチ数列、フラクタルなどの数学的原理は、方向性のないランダムな進化の産物ではなく、正確な目的で生命体の遺伝子に組み込まれた精巧な設計の結果であることを示している。そうでなければ、ヒマワリやワラビのような植物が数学が得意で、黄金角に沿って種をつけ、フラクタル構造に沿って葉を出したと信じなければならないだろう。
4.福音 への招待
「あなたの指のわざである天を見、あなたが整えられた月や星を見ますのに、
人とは、何者なのでしょう。あなたがこれを心に留められるとは。人の子とは、何者なのでしょう。あなたがこれを顧みられるとは。
あなたは、人を、神よりいくらか劣るものとし、これに栄光と誉れの冠をかぶらせました。
あなたの御手の多くのわざを人に治めさせ、万物を彼の足の下に置かれました。
すべて、羊も牛も、また、野の獣も、
空の鳥、海の魚、海路を通うものも。
私たちの主、主よ。あなたの御名は全地にわたり、なんと力強いことでしょう。」(詩篇8:3-9)
上記の聖書箇所で、詩人は夜空に広がる天と星々を見上げながら、万物を創造された主なる神を賛美し、同時に「人間とはいったい何者であるのか」と、自らの存在の意味を思い巡らせています。この本を通して私たちは、広大な宇宙、地球、そしてそこに存在するあらゆる生命体が、決して偶然の結果ではなく、すべてが創造主である神様の深い知恵と力によって造られたことをはっきりと学んできました。
では、なぜ神様は巨大な宇宙を創造し、この地球に命を与え、私たち人間を生かしておられるのでしょうか。聖書はその創造の目的を明確に示しています。それは、神様が私たち人間との永遠の交わりを望んでおられるからです。聖書には「地の 果てのすべての 者よ。 わたしを 仰ぎ 見て 救われよ。 わたしが 神である。 ほかにはいない。」(イザヤ書45章22節)と記されています。この言葉からも分かるように、神様は私たちに救いを与え、永遠の御国の市民として共に生きることを望んでおられます。
しかし、多くの人々は、この世の人生が終わればすべてが終わると考え、地上の事柄に心を奪われてしまっています。そのため、神様が与えてくださる救いの機会を逃しているのが現実です。イエス・キリストはこの現実について、「大宴会のたとえ」(ルカの福音書14章15節~24節)や「婚宴のたとえ」(マタイの福音書22章1節~14節)において、詳しく語っておられます。
これらのたとえ話では、神様が人々を天の御国への祝宴に招いておられるにもかかわらず、招かれた人々がさまざまな理由を挙げてその招待を拒む様子が描かれています。「私は畑を買ったので行けません」「牛を試しに行かなければなりません」「結婚したばかりなので都合がつきません」などと、それぞれが自分の都合を優先し、せっかくの招きを断ってしまうのです。
囲碁を指す人でさえ、常に数手先を見据えて最善の手を打とうとします。しかし、永遠の運命を決する重大な招きが、ただこの地上で生きている間にしか与えられないにもかかわらず、多くの人がそれを軽んじ、世のことに忙殺されて神様の招きを退けているのは、なんと惜しいことでしょう。
この章では、あなたを天国へと導く「福音」についてご紹介したいと思います。しかし、まだ神様の存在そのものを信じることが難しいと感じている方もいらっしゃるかもしれません。そこで、まず「神様の存在を信じることは合理的である」とする考え方を示す「パスカルの賭け(Pascal's Wager)」についてご説明させていただきます。
哲学者であり、物理学者・数学者、さらには神学者でもあったブレーズ・パスカルは、彼の著書『パンセ』の中で、人々を真理へと導くためにキリスト教の教義を説明し、その合理性を弁証しようと試みました。彼は「人間は考える葦である」という有名な言葉を残しています。広大な宇宙の中で、人間はまるでひとつの点のように小さく、儚い存在にすぎません。しかし、パスカルは次のように語っています。「空間は私を包み込み、一つの点として私を押し潰すかもしれない。しかし、私の思考はこの広大な宇宙をも包み込むことができる」と。
パスカルは、「人間の真の喜びは、創造主である神を知ることにある」と述べ、さらに「パスカルの賭け」と呼ばれる次のような考え方を提示しました。
神が存在すること | 神が存在しないこと | |
神の存在を信じる | 永遠の喜び (天国) | 正しく世界を生きる |
神の存在を信じない | 永遠の苦しみ (地獄) | 無神論者として生きる |
表4.1.パスカルの賭け
これまで、私たちは天地万物が決して偶然によるものではなく、創造主である神様の御手によって造られたことを学んできました。ですから、私たちの前には二つの選択肢があります。一つは、神様を信じて永遠の神の国、すなわち天国への道を選ぶこと。もう一つは、神様を信じないまま、永遠の滅び、すなわち地獄へと向かう道を選ぶことです。この選択において、私たちは誰もが前者を選ぶべきではないでしょうか。
天国や地獄の存在について、ある方にはそれが神話のように感じられるかもしれません。しかし、聖書はこれらが紛れもない現実であることを、はっきりと証言しています。たとえば、『コリント人への手紙 第二』12章2〜4節には、次のように記されています。
「私はキリストにあるひとりの人を知っています。この人は十四年前に――肉体のままであったか、私は知りません。肉体を離れてであったか、それも知りません。神はご存じです。――第三の天にまで引き上げられました。
私はこの人が、――それが肉体のままであったか、肉体を離れてであったかは知りません。神はご存じです。――
パラダイスに引き上げられて、人間には語ることを許されていない、口に出すことのできないことばを聞いたことを知っています。」
この体験を語ったのは、かつてキリスト教徒を激しく迫害していた使徒パウロです。彼はダマスコへの途上で復活したイエス・キリストと出会い、回心しました。パウロの証言によれば、天国の楽園は「第三の天」にあり、そこは肉体では行けず、霊によってのみ至ることができる場所であるとされています。また、パウロは三次元の世界に生きる私たちには理解しがたい、神の奥義を体験し、人間には語ることが許されていない大いなる秘密が存在することを知りました。
神様は、すべての人が永遠の命を得て、天国に住むことを心から望んでおられます。そのため、永遠の昔から神様と共におられた御子イエス・キリストを、この地に送られたのです。イエス・キリストがこの地に来られたことは、歴史的にも確かな事実です。実際に、人類の歴史は「キリスト以前(B.C.: Before Christ)」と「主の年(A.D.: Anno Domini – Latin for in the year of our LORD)」によって区分されています。
イエスは地上で、人間には決して成し得ない数々の奇跡を行われました。天から来られた方のみが語ることのできる教えを説き、死んだラザロをよみがえらせ、生まれつき目の見えなかった人を癒し、五つのパンと二匹の魚で五千人を養い、風と波を鎮め、水の上を歩くなど、驚くべき御業を次々と示されました。悪霊でさえイエスの御前では震え上がり、命令に従わざるを得ませんでした。これらの奇跡はすべて、イエスこそが命の主であり、天地万物の主であり、神から遣わされた救い主であることの揺るぎない証拠です。
イエス・キリストがこの世に来られた最も大切な目的は、私たちを罪から救うためでした。「罪」とは、神を信じず、自分中心に生きることから生じる不信仰と、それによって現れる行動――嘘や詐欺、偶像崇拝、貪欲、傲慢、姦淫、怒り、嫉妬などです。この罪によって、神と人との関係は断絶されてしまいました。
しかし、神はご自身に似せて造られた私たち人間を深く愛し、その罪の問題を解決するためにイエス・キリストをこの世に遣わされたのです。罪のない義なるイエス・キリストは、私たちすべての罪の代価として「死」を、十字架において身代わりに負ってくださいました。そして、三日目に復活され、死に打ち勝たれました。その後、弟子たちの見守る中、天に昇られ、今も永遠の主として生きておられます。
ですから、誰であっても、自分が神の前で「罪人」であることを認め、このイエス・キリストを、自らの罪のために十字架にかかって死に、復活された救い主として信じ受け入れるなら、その人の罪は赦され、神の子として新しく生まれ変わり、永遠の命が与えられます。神の国の民とされるのです。これこそが「福音」-神がすべての人に与えておられる計り知れない恵みの知らせです。
永遠の命は、人間の努力や善行によって得られるものではありません。これは、ただ神の恵みによって、信仰を通して与えられる「贈り物」なのです。
「でも、こんなにも素晴らしい話、信じられない」と感じる方もいるでしょう。しかし、これはまぎれもない真実であり、神からあなたへの招きです。福音は、罪によって疎遠になった神との関係を回復し、永遠に続く交わりへと導く道です。
もしあなたが、イエス・キリストを救い主として受け入れたいと願うなら、ぜひ主日にお近くの教会を訪ねてください。そして、牧師にその思いをお伝えください。きっと、祈りを共にし、イエス様がなされたこと、そして信仰の歩みについて親切に教えてくれることでしょう。以下は、イエス様を心に迎え入れるための「祈りの例」です。
《祈りの例》
「天の父なる神様、私は罪人です。今まで、どこから来て、どこへ行くのかも分からず、迷いながら生きてきました。自分の欲望と肉の思いに従い、あなたを悲しませる多くの罪を犯してきました。けれども、あなたはそんな私をなおも愛し、御子イエス・キリストをこの地に送ってくださいました。イエス様は、私のすべての罪を代わりに背負い、十字架で死んでくださり、三日目によみがえられたことを私は信じます。今、心の扉を開き、イエス・キリストを私の救い主としてお迎えします。どうか、私をお受け入れくださり、私の罪を赦し、永遠の命をお与えください。イエス・キリストのお名前によって祈ります。アーメン。」
イエス・キリストを救い主として心に迎え入れたなら、これまで抱えていた不安や恐れは、天からの平安によって包まれていくことでしょう。
イエスがあなたの主であるなら、もはや何を恐れる必要があるでしょうか。あなたの世界観は変わり、人生の優先順位も新しくされていくはずです。地上のものだけを追い求め、やがて虚しく終わる人生ではなく、永遠に価値ある「天のもの」を目指す、希望に満ちた新しい人生が始まります。
どうかこの主日が、あなたにとって「新しい霊的な世界」への扉となり、人生で最も価値ある一日となることを、心からお祈りしています。
「主イエスを信じなさい。そうすれば、あなたもあなたの家族も救われます。」(使徒の働き16:31)
謝辞
この本の推薦文を書いてくださり、また初稿を最初から最後まで丁寧に読み、細心の注意を払って修正し、必要な内容を補ってくださったCalifornia Bridge韓人教会の朴環哲宣教師に、心より深く感謝申し上げます。
さらに、聖書と天文学に関する多くの対話を通して、この本の執筆に大きな動機を与えてくださったKim Yong-Cheol牧師、Kim Jong-Kug牧師、Kim Kyoung-A宣教師、Kim Hyun-A姉妹にも感謝の意を表します。
また、原稿を読んで貴重なフィードバックをくださったCharlottesville BLOO-gene韓人教会のIm Jun-Sup牧師、Arcturus TherapeuticsのChoi Kyoung-Joo博士、KRICTのPark Chi-Hoon博士にも感謝いたします。加えて、画像作業を手伝ってくれた長男サミュエルと次男ダニエルにも感謝の気持ちを伝えます。
最後に、この日本語訳の草稿を丹念に検討し、文法の誤りを修正し、文章表現を洗練してくださった朴環哲宣教師、Choi Kyoung-Joo博士、Yoon Chan-Yang姉妹に、重ねて深く感謝申し上げます。
すべての栄光を神に!
推薦の声
この本の著者である金東燦博士と初めてお会いしたのは、今から20年前、日本の神奈川県大和市でのことでした。私が開拓教会である大和バプテスト教会を牧会して4年ほど経った頃, 金博士はアメリカから日本の宇宙航空研究開発機構(JAXA)に滞在され、天文学の研究に従事しておられました。ある主日、神様を礼拝するために、偶然にも私の教会を訪ねてくださったのです。
礼拝後のご挨拶を交わす中で、金博士がクリスチャンであり、しかも天文学の博士でいらっしゃることを知りました。それまで私は天文学に関してほとんど知識がなく、縁遠い分野だと感じておりましたが、博士の信仰と科学に対する真摯な姿勢に強く心を惹かれ、思わず「天文学者として教会でセミナーをしていただけませんか」とお願いしました。
当時、私たちの教会は開拓段階で、外部から講師を招く機会などありませんでした。けれども、新たなセミナーを開けば、聖徒たちにとって貴重で新鮮な経験となるだろうと思い、軽い気持ちでお願いしたのを覚えています。金博士は快く引き受けてくださり、実はこれまでもいくつかの教会で同様のセミナーを行われた経験があることを教えてくださいました。
私たちは日程を調整し、簡単なポスターを作って市内の掲示板に貼って告知しました。そして、「天文学的観点からの信仰と創造論」というテーマで一日セミナーを開催することになったのです。場所は、商店街の建物の2階にある小さな礼拝堂でしたが、教会の聖徒たちに加え、数名の日本人の方々も参加してくださいました。金博士は誠心誠意、心を込めてセミナーを導いてくださいました。参加者の多くが深い感銘を受けましたが、中でも特に私自身にとっては、非常に大きな感動をもたらすものでした。その感動を通じて「この話をもっと多くの人に届けたい」との思いが心の中に強く芽生え、ずっとその機会を探し続けてきました。
そんな折、神奈川県内のバプテスト教会の牧師たちの定期的な集まりで知り合った、平塚市のバプテスト教会の梶井義郎牧師が、毎年年末に市内の大きな劇場や講堂を借りて、多くの市民を招き「市民クリスマス公演」を開催されていることを知りました。私はすぐに金博士のセミナーを紹介し、その年の年末には、金博士が再びアメリカから来日され、平塚市民の前で「天文学的観点からの創造論」をテーマにセミナーを行っていただくことになったのです。
このセミナーを通して私が受けた感動と悟りは、私自身の人生を新たに見つめ直すきっかけとなりました。それは、これまで当然のように見過ごしてきた自然や周囲のすべての存在への認識を一変させるものでした。生命の一つひとつが、いかに貴重で尊いものであるのかを改めて実感し、私の目は新たに開かれたのです。それは、私が10代の終わりに初めて主と出会い、魂が新たに生まれ変わった時の感動とはまた異なる次元の、自然と生命に対する霊的な目覚めの瞬間でした。
この広大な宇宙に無数の銀河が存在し、さらにその中に無数の惑星がある中で、地球というこの小さな惑星が特別に造られ、生命が存在できるような環境として創造主によって緻密に整えられ、守られ、管理されている。その事実に、私はまさに驚異としか言いようがありません。そして、地球上のすべての生命体が、どれほどかけがえのない、尊い存在であることか。道を歩くとき何気なく踏みしめる草の一本、地面を這う小さな蟻一匹さえも、尊く大切な命として見えるようになりました。そして、万物の霊長として造られた人間である私は、一体何者なのか。なぜこれほどまでに尊い存在として創造され、生かされているのか。私は深く思いを馳せるようになりました。
金博士のセミナーを通して得たこの感動と気づきは、学ぶ者として生涯を歩みたいと願ってきた私の人生を揺るがし、これまでのどんな学びや経験からも得られなかったほどの満足と喜びを与えてくれました。それは単なる現代天文学の成果や知識の紹介ではなく、私たちの命そのものの根源にまで導いてくださる、偉大なる神への畏敬の念と、生命の価値、その崇高さを見つめる眼差しを育ててくれるものでした。
私にとって今なお深い余韻として残る金博士のセミナーは、20年前に限られた時間の中で開催されたものでしたが、この度出版された本には、当時のセミナー内容がより詳しく記され、博士が長年にわたり研究・収集してこられた科学的資料も加えられています。そして、博士自身の信仰をもって、読者に創造主なる神の人類に対する永遠の救いの計画、すなわち福音を紹介し受け入れるようにと語りかけています。
アブラハム・J・ヘーシェルは、著書『人を捜す神』、第4章「驚異」の中で、「現代人は文明の発展に伴い、すべてを科学で説明できると考え、あらゆる事象を当たり前のものとして捉える傾向が強くなっているため、驚異の感覚が失われつつある」と指摘しています。そして、「自然や存在に対する驚異こそが、人生に対する感動と感謝を呼び起こし、その崇高な驚異が信仰の第一歩となる」と述べています。さらには、「私たちの幸せの始まりは、驚異のない人生は生きる価値がないことを悟ることであり、私たちに欠けているのは信じようとする意志ではなく、驚異を感じようとする意志である」と語っています。
どうかこの本を手に取られるすべての方々が、私が金博士のセミナーで味わったような感動と気づきと同じく、創造主である神への畏敬の念が新たにされ、神が創造された万物に対する驚異の目が開かれることを心より願っております。そして、皆さまが神の偉大で驚くべき愛を深く悟る機会となりますことを祈りつつ、推薦の言葉とさせていただきます。
朴環哲、大和バプテスト教会牧師、日本宣教師(2000-2006)
画像出典
1. 宇宙の創造
Fig. 1.1: NASA/JPL, Fig. 1.2: Hubble Heritage Team, Fig. 1.3: R. Hurt/JPL-Caltech/NASA, Fig. 1.4: Hubble/NASA/ESA, Fig. 1.5: Wikipedia/R. Powell, Fig. 1.6: Wikimedia/D. Leinweber, Fig. 1.7: NASA/CXC/M. Weiss(left), NASA/D. Berry (right), Fig. 1.8: Stellarium, Fig. 1.9: Physics Forums, Fig. 1.10: NASA/JPL-Caltech (left), A. Sarangi, 2018, SSR, 214, 63 (right), Fig. 1.11: Wikimedia/ALMA (ESO/NAOJ/NRAO) (left), T. Müller (HdA/MPIA)/G. Perotti (The MINDS collaboration)/M. Benisty (right), Fig. 1.12: TASA Graphic Arts, Inc., Fig. 1.14: Jon Therkildsen, Fig. 1.15: www.neot-kedumim.org.il
2. 神様の傑作 地球
Fig. 2.1: R. Narasimha, Fig. 2.3: NASA, Fig. 2.4: NASA/Goddard/Aaron Kaase, Fig. 2.6: Wikimedia, Fig. 2.7: Linda Martel, Fig. 2.8: Wikimedia, Fig. 2.9: NASA/ESA/H. Weaver & E. Smith (left), NASA/HST Comet Team (right), Fig. 2.10: Wikimedia/M. Bitton, Fig. 2.11: Wikimedia/John Garrett, Fig. 2.12: UK Foreign and Commonwealth Office, Fig. 2.13: Wikipedia, Fig. 2.16: Wikipedia/G. Taylor, Fig. 2.17: NASA/Caltech
3. 創造か進化か?
Fig. 3.1: Wikipedia/Yassine Mrabet, Fig. 3.2: OpenEd/Christine Miller , Fig. 3.3: Wikipedia/LadyofHats, Fig. 3.4: Wikipedia/Messer Woland & Szczepan (left), Wikipedia/LadyofHats (right), Fig. 3.5: J.E. Duncan & S.B. Goldstein, Fig. 3.6: Wikipedia/Fiona 126, Fig. 3.7: NASA, Fig. 3.8: R. Cui, Fig. 3.9: Wikipedia/Ansgar Walk, Fig. 3.10: The Whisker Chronicles, Fig. 3.11: Encyclopedia Britanica Inc., Fig. 3.12: Wikipedia, Fig. 3.13: Wikipedia/LadyofHats, Fig. 3.14: Wikipedia/J.J. Corneveaux, Fig. 3.15: Smithsonian Institution, Fig. 3.17: NRAO/AUI/NSF (left), Wikipedia/Colby Gutierrez-Kraybill (right), Fig. 3.18: Wikipedia/MikeRun, Fig. 3.20 - Fig. 3.23: Shueisha, Inc./Obara Takuya, Fig. 3.24: Wikipedia/Pinakpani, Fig. 3.25: Wikipedia/Dicklyon, Fig. 3.26: Wikipedia/Stannered (1st img), Dicklyon (2nd img), Morn the Gom (3rd img), Eequor (4th img), Fig. 3.27: M. Kitazawa/J. Plant Res., Fig. 3.28: S.R. Rahaman, Fig. 3.30: Jill Britton (pineapple), Fig. 3.32: Wikipedia/Farry (left), Wikimedia/Ivar Leidus (right).
参考文献
1. 宇宙の創造
제자원 (2002), Oxford Bible Encyclopedia, Bible Textbook Co., Genesis Chap. 1-11.
Another universe existed before Big Bang? 우주먼지의 현자타임즈, 2/24/2024, https://www.youtube.com/watch?v=RckLkaVzFe0
A Big Ring on The Sky: AAS 243rd Press conference. Alexia M. Lopez, 1/11/2024, https://www.youtube.com/watch?v=fwRJGaIcX6A
Bogdan, A., et al. (2024), “Evidence for heavy-seed origin of early supermassive black holes from a z ≈ 10 X-ray quasar”, Nature Astronomy, 8, 126.
Bonanno, A., & Fröhlich, H.-E. (2015), “A Bayesian estimation of the helioseismic solar age”, Astronomy & Astrophysics, 580, A130.
Karim, M. T., & Mamajek, E. E. (2017), “Revised geometric estimates of the North Galactic Pole and the sun's height above the Galactic mid-plane”, MNRAS, 465, 472.
Lopez, A. M., et al. (2022), “Giant Arc on the sky”, MNRAS, 516, 1557.
Lopez, A. M., Clowes, R. G., & Williger, G. M. (2024), “A Big Ring on the Sky”, JCAP, 07, 55.
Lyra, W., et al. (2023), “An Analytical Theory for the Growth from Planetesimals to Planets by Polydisperse Pebble Accretion”, The Astrophysical Journal, 946, 60.
Penrose, R. (2016), The Emperor’s New Mind, Oxford University Press, Oxford, United Kingdom.
Perotti, G., et al. (2023), “Water in the terrestrial planet-forming zone of the PDS 70 disk”, Nature, 620, 516.
Sandor, Zs., et al. (2024), “Planetesimal and planet formation in transient dust traps”, Astronomy & Astrophysics, in press.
Schiller, M., et al. (2020), “Iron isotope evidence for very rapid accretion and differentiation of the proto-earth”, Science Advances, 6, 7.
Tonelli, G. (2019), Genesis: The story of how everything began, Farrah, Straus and Giroux, New York, pp 19-44
Tryon, E. P. (1973), “Is the Universe a vacuum fluctuation”, Nature, 246, 396.
Vorobyov, E. I., et al. (2024), “Dust growth and pebble formation in the initial stages of protoplanetary disk evolution”, Astronomy & Astrophysics, 683, A202.
Yi, S., et al. (2001), “Toward Better Age Estimates for Stellar Populations: The Y2 Isochrones for Solar Mixture”, The Astrophysical Journal Supplement Series, 136, 417.
2. 神様の傑作 地球
Comins, N. F. (1993), What If the Moon Didn't Exist? HarperCollins Publishers Inc., New York, NY.
Gonzalez, G. & Richards, J. W. (2004), The privileged planet: How Our Place in the Cosmos Is Designed for Discovery, Regnery Publishing, Inc.
Lineweaver, C. H., et al. (2004), “The Galactic Habitable Zone and the Age Distribution of Complex Life in the Milky Way”, Science, 303 (5654), 59.
Lüthi, D. et al. (2008), “High-resolution carbon dioxide concentration record 650,000 - 800,000 years before present”, Nature, 453, 379.
Narasimha, R., et al. (2023), “Making Habitable Worlds: Planets Versus Megastructures”, arXiv:2309.06562.
OpenAI. (2024), ChatGPT (4o) [Large language model], https://chatgpt.com
Ward, Peter D. & Brownlee, Donald (2000), Rare Earth: Why Complex Life is Uncommon in the Universe, Copernicus Books (Springer Verlag).
3. 創造か進化か?
Abelson, P. H. (1966), “Chemical Events on the Primitive Earth”, Proc Nat Acad Sci, 55, 1365.
Behe, M. J. (2006). Darwin's black box: The biochemical challenge to evolution. Free Press.
Behe, M. J. (2020). Darwin devolves: The new science about DNA that challenges evolution. HarperOne.
Bernhardt, H. S. (2012), “The RNA world hypothesis: the worst theory of the early evolution of life (except for all the others)”, Biology Direct, 7, Article number: 23.
Chyba, C. F., & Sagan, C. (1992), “Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: An inventory for the origins of life”. Nature, 355, 125.
Cui, R., “The transcription network in skin tanning: from p53 to microphthalmia”, https://www.abcam.com/index.html?pageconfig=resource&rid=11180&pid=10026
Dembski, W. A., & Ewert, W. (2023). The design inference: Eliminating chance through small probabilities. Discovery Institute.
Danielson, M. (2020), “Simultaneous Determination of L- and D-Amino Acids in Proteins”, Foods, 9 (3), 309.
Fabre, J.-H. (2015), The Mason -Bees (Perfect Library), CreateSpace Independent Publishing Platform.
Higgins, M. (2014), “Bear evolution 101”, The Whisker Chronicles, https://thewhiskerchronicles.com/2014/01/03/bear-evolution-101/
Kasting, J. F. (1993). “Earth's Early Atmosphere.” Science, 259(5097), 920.
Maslin, M. (2016), “Forty years of linking orbits to ice ages”, Nature, 540 (7632), 208.
Miller, S. L. (1953), “A Production of Amino Acids under Possible Primitive Earth Conditions”, Science, 117, 528
Mumma, M. M., et al. (1996), “Detection of Abundant Ethane and Methane, Along with Carbon Monoxide and Water, in Comet C/1996 B2 Hyakutake: Evidence for Interstellar Origin”, Science, 272 (5266), 1310.
OpenAI. (2024), ChatGPT (4o) [Large language model], https://chatgpt.com
Park, Chi Hoon (2024), “Stop codon points to GOD”, Proceedings of the 20th Anniversary KRAID Symposium
Pinto, J. P., Gladstone, G. R., & Yung, Y. L. (1980), “Photochemical Production of Formaldehyde in Earth’s Primitive Atmosphere”, Science, 210, 183.
Pinto, O. H., et al. (2022), “A Survey of CO, CO2, and H2O in Comets and Centaurs”, Planet. Sci. J., 3, 247.
Russo, D., et al. (2016), “Emerging trends and a comet taxonomy based on the volatile chemistry measured in thirty comets with high resolution infrared spectroscopy between 1997 and 2013”, Icarus, 278, 301.
Sanjuán, R., Moya, A., & Elena, S. F. (2004), “The distribution of fitness effects caused by single-nucleotide substitutions in an RNA virus”, Proc Natl Acad Sci, 101(22), 8396.
Trail, D., et al. (2011), “The oxidation state of Hadean magmas and implications for early Earth’s atmosphere”, Nature, 480, 79.
Urey, H. C. (1952). “On the Early Chemical History of the Earth and the Origin of Life.” Proc Natl Acad Sci, 38(4), 351.
Wikipedia, Mutation (Distribution of fitness effects).
Wikipedia, Visual phototransduction.
Yang, P.-K. (2016), “How does Planck’s constant influence the macroscopic world?”, Eur. J. Phys., 37, 055406.
Zahnle, K. J. (1986), “Photochemistry of methane and the formation of hydrocyanic acid (HCN) in the Earth’s early atmosphere”, J. Geophys Res, 91, 2819.
著者紹介
金東燦博士は、韓国ソウルの延世大学で天文学を専攻し、ハワイ大学マノア校で天文学博士号を取得。 博士課程修了後、NASAジェット推進研究所/カリフォルニア工科大学、ソウル大学、バージニア大学など複数の機関で天文学研究を行いました。
金博士の研究テーマは、発光赤外線銀河(LIRG)、超光赤外銀河(ULIRG)、クエーサー、反動超巨大ブラックホールです。
現在、米国バージニア州シャーロッツビルにある国立電波天文台に勤務している。
この本は[DIVINE GENESIS: Exploring Creation through Astronomy and Biology]というタイトルでアメリカのAmazonで出版された本の日本語訳本です。The PDF version of this book, along with translations in multiple languages, can be downloaded from divine-genesis.org.