천문학과 생물학적 관점에서본
창조론
서문
이 책은 우주의 창조, 하나님의 걸작품 지구, 그리고 생명의 기원에 대한 논의를 통해 창조론과 진화론을 탐구하기 위해 쓰여졌다.
첫째, 우주의 창조를 논하기 위해 먼저 우주의 광대함과 그 층상구조에 대해 간략하게 소개한 후, 최신 천문학적 연구를 통해 밝혀진 우주의 기원과 우주의 운명에 대해 살펴볼 것이다. 그런 다음, 성경의 우주 창조 기록이 천문학적 관측과 일치하는지, 지구와 태양의 창조, 일부 창조론자들이 주장하는 지구 나이가 6천 년인지, 그리고 미세 조정된 우주에 대해서도 살펴보고자 한다.
둘째, 하나님의 걸작품 지구에서는 지구가 우주에서 얼마나 특별한 행성인지를 10가지 주요 사실을 통해 살펴보고, 이를 바탕으로 지구가 단순한 우연의 산물인지 아니면 생명체의 생존을 위해 특별하게 창조된 행성인지에 대해 알아볼 것이다.
셋째, 생명체의 기원에 관한 다양한 논의를 통해 진화론과 창조론의 타당성을 검토해 보려고 한다. 그리고, 생명체의 환경 적응에 대한 논의를 통하여 다윈의 이론이 진화론으로 불려야 하는지 환경에 대한 유전적 적응론으로 불려야 하는지, 인간이 유인원에서 진화했는지, 지적 설계론은 무엇인지에 대해서도 알아볼 것이다. 아울러 입자물리학, 외계인의 존재 가능성, 동물의 본능, 자연계에서 발견되는 수학적 원리를 통해 창조론을 살펴볼것이다.
마지막으로, 여러분을 믿음으로 초대하고자 한다. 현대 사회는 물질만능주의와 수많은 미디어의 영향으로 인해 정신적 가치가 왜곡되고 인간 본연의 숭고함과 가치 있는 삶의 기준이 흐려지고 있다. 이러한 시대 속에서 "나는 어디에서 왔으며, 어디로 가고 있는가?"라는 근본적인 질문을 던져 본 적이 있다면 그 해답으로 영생으로 인도하는 복음을 소개하고자 한다.
이 책을 통해 독자들이 생명의 기원이 어디에서 왔는지, 지구상의 모든 생명체가 얼마나 소중한 존재인지 다시금 생각해 보는 기회가 되기를 바란다. 그리고, 이 모든 것을 창조하시고 보존하시는 창조주에 대한 경외감을 가지게 되는 계기가 되길 소망한다.
|
|

1. 우주의 창조
어린 시절 시골이나 높은 산에서 캠핑하며 드넓은 밤하늘에 반짝이는 수많은 별을 바라보거나 어두운 하늘을 우아하게 가로지르는 별똥별을 보며 감탄했던 기억이 있을 것이다. 이런 경험은 종종 우리를 우주에 대한 경외감으로 가득 차게 하며 여러 의문을 품게 한다. 우주에는 얼마나 많은 별이 있을까? 지구 너머에 생명체가 존재할 수 있을까? 우주는 어떻게 시작되었고, 어떻게 끝날까? 누가 이 모든 것을 창조했을까? 등등 여러 의문을 품게 할 것이다.
이 장에서는 천문학적 관점과 성경적 관점에서 우주의 기원을 살펴보고자 한다. 이 두 가지 관점을 비교함으로써 성경 창세기의 창조 기록이 천문학적 관측과 일치하는지 알아볼 것이다. 또한 지구와 태양 중 어느 것이 먼저 창조되었는지, 지구의 나이가 6,000년인지, 미세 조정된 우주에 대해서도 살펴볼 보고자 한다.
a. 우주의 계층적 구조
우주의 기원에 대해 논의하기 위해 먼저 우주의 계층적 구조를 살펴보기로 하자. 태양계부터 시작하여 은하, 외부 은하, 은하단, 초은하단, 초은하단 Complex로 나아가 보겠다.
i. 태양계
‘태양계’란 태양으로 불리는 별(항성)과 그 주위를 공전하는 8개 행성, 화성과 목성 사이의 소행성대, 명왕성이 포함된 카이퍼 벨트, 그리고 맨 마지막에 위치한 오르트 구름으로 구성되어 있다. 여기서 별이란 핵융합 반응에 의해 자기 스스로 빛을 내는 천체를 말하며, 행성이란 스스로 빛을 내지 못하지만 별에서 오는 빛을 반사하여 밝게 빛나는 천체를 말한다. 행성은 대부분 암석으로 이루어진 지구형 행성(Terrestrial planets)과 대부분 가스로 이루어진 목성형 행성(Jovian planets)으로 나눌 수 있다. 우리 지구는 태양으로부터 3번째 위치한 지구형 행성이다.
지구에서 달까지의 거리는 38만 4천km이고 시속 1,000km의 비행기로 가면 16일이 걸린다. 지구로부터 태양까지의 거리는 약 1억 5천만km이며 비행기로 가면 17년이 걸린다. 천문학에서는 지구에서 태양까지의 거리를 1천문단위로 정의한다. 이 천문단위를 쓰면 맨 바깥에 위치한 행성인 해왕성까지는 30천문단위, 카이퍼 벨트는 30~50천문단위, 오르트 구름까지는 2,000~200,000천문단위가 된다. 우리가 관측하는 혜성들은 단주기 혜성과 장주기 혜성이 있다. 단주기 혜성은 카이퍼 벨트의 물체가 섭동을 받아 태양의 중력에 이끌려 오는 것이고 장주기 혜성은 오르트 구름의 물체가 섭동을 받아 태양의 중력에 이끌려 오는 것이다. 혜성들이 그렇게 먼 거리에서 태양을 방문하기에 이심률이 큰 타원 궤도를 가지게 된다.
빛의 속도로 달리면 지구로부터 태양까지는 8.3분, 해왕성까지는 4시간, 태양계 끝까지는 9개월 반 또는 0.79광년(1광년 = 빛이 1년 동안 달리는 거리)이 걸린다. 시속 1,000km의 비행기로 가면 약 85만 년이 걸린다. 태양은 지구의 109배 크기, 질량은 333,000배이며 자전 주기는 약 25일이다.
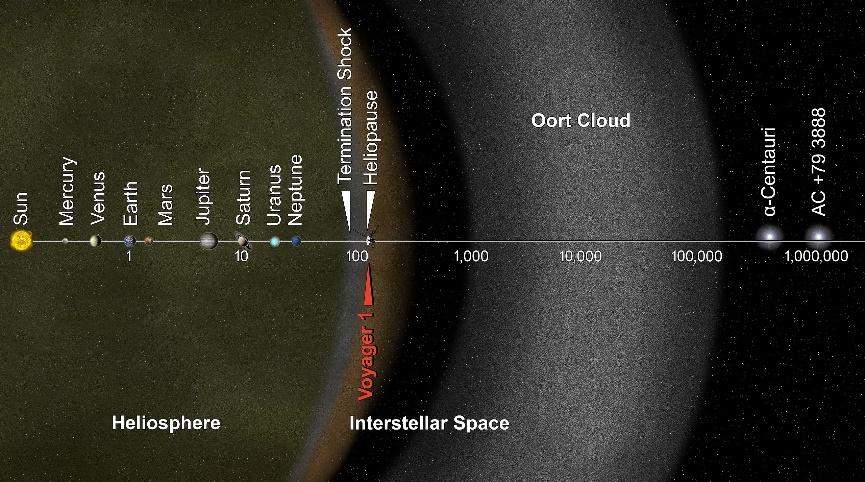
그림 1.1. 오르트 구름을 포함한 태양계
ii. 별의 세계(항성계)
오르트 구름을 벗어나면 이제 별의 세계로 들어간다. 지구에서 가장 가까운 별은 ‘프록시마 센타우리’라는 별이고 크기는 태양의 14%, 질량은 태양의 12%, 거리는 약 4.2광년 떨어져 있다. 시속 1,000km 비행기로 간다면 약 457만 년이 걸린다.
우리가 밤하늘에 보는 반짝이는 별들은 자세히 보면 여러 색깔의 별들이 있다는 것을 알게 된다. 별의 색깔은 별 표면의 온도에 따라 달라지는데 온도가 낮으면 붉은색을 온도가 높으면 청백색을 띠게 된다. 대장간에서 무쇠를 달구면 처음에는 붉은색을 띠다가 온도가 올라가면 노란색으로 그리고 온도가 더 올라가면 청백색으로 변하는 것과 같은 이치이다. 오리온자리의 알파성인 베텔게우스는 붉은색을, 우리 태양은 노란색, 그리고 사냥개자리의 알파성인 시리우스는 청백색을 띠고 있다. 이런 차이를 가져오는 가장 근본적인 요인은 별의 질량이며 별의 진화 과정을 결정하는 것도 별의 질량이다.

그림 1.2. 다양한 색상을 가지고 있는 별들
별의 질량은 핵융합 속도를 결정하고, 이는 다시 별의 광도와 수명을 결정한다. 질량이 큰 별은 질량이 작은 별보다 연료를 더 빨리 소모한다. 별이 핵연료를 다 쓰고 도달하는 마지막 단계는 백색왜성, 중성자별, 또는 블랙홀이다. 별이 적색거성 단계를 거친 후 중심부 질량이 태양의 약 1.4배 이하인 별들은 중심에서의 헬륨 핵융합 반응 이후 계속 수축해서 백색왜성이 되며 껍질은 팽창하여 행성상 성운이 된다. 중심부 질량이 태양의 약 1.4배 이상이고 3배 이하인 별들은 중심부에서 양성자가 전자와 결합하여 중성자별이 되고, 중심부 질량이 태양의 3배 이상인 별들은 강력한 중력으로 인해 중성자별을 거쳐 블랙홀이 된다. 중성자별과 블랙홀의 껍질은 격렬하게 폭발하여 초신성이 되며, 폭발한 초신성의 잔해가 모여 중력적으로 수축하면 다시 새로운 별들이 만들어진다.
현재 우리가 육안으로 볼 수 있는 별의 개수는 도시에서는 백 개 정도, 시골 오지에서는 천 개 정도 볼 수가 있다. 이러한 별들은 대부분 지구로부터 약 50광년 이내에 있는 아주 가까운 별들이다.
iii. 우리은하
우리은하는 막대 나선은하로 분류되며 약 2천억-4천억 개 정도의 별이 모여 있다. 크기는 지름이 약 10만 광년이고 두께는 약 1천 광년이다. 태양은 은하 중심으로부터 약 2만 6천 광년 떨어져 있으며 약 2억 1천만 년 주기로 은하 중심을 공전하고 있다. 태양계는 페르세우스 팔과 방패-센타우루스자리 팔 사이에 자리 잡고 있으며 은하면에서부터 약 60광년 위쪽에 있어 어느 방향을 보더라도 우주를 잘 관측할 수 있는 전망 좋은 자리에 있다.

그림 1.3. 우리은하
iv. 은하, 은하단 및 초은하단
우리은하를 벗어나면 은하들의 세계가 펼쳐진다. 우리에게 가장 가까운 은하는 안드로메다은하이며 거리는 약 250만 광년이 된다. 육안으로 관측이 가능하며 우리은하와 비슷하게 생겼다. 안드로메다은하는 초당 약 110km의 속도로 우리은하로 접근하며 약 40억 년 후에는 우리은하와 충돌할 것으로 예상된다. 은하를 시각적 형태로 분류하면 나선은하, 타원은하, 그리고 불규칙 은하로 나눌 수 있다. 두 개의 나선은하가 충돌하면 밝은 적외선 은하(Luminous infrared galaxies, Ultraluminous infrared galaxies)를 거쳐 타원은하가 만들어진다.

그림 1.4. 나선 은하, 타원 은하, 불규칙 은하
50개 미만의 은하들이 중력적으로 모여 있으면 은하군(Group of galaxies)이라 부르고 수백 내지 수천 개의 은하가 모여 있으면 은하단(Cluster of galaxies)이라 부른다. 우리은하와 안드로메다은하 등 가까운 40여 개의 은하들은 국부 은하군(Local Group)에 속하며, 국부 은하군과 처녀자리 은하단(Virgo Cluster)은 처녀자리 초은하단(Virgo Supercluster)에 속하고, 처녀자리 초은하단은 그보다 더 큰 라니아케아 초은하단(Laniakea Supercluster)에 속한다.
은하 필라멘트 또는 초은하단 사슬이라고도 하는 초은하단 Complex는 은하, 가스, 암흑 물질의 광대한 네트워크로 서로 연결된 수많은 은하단으로 구성된 우주의 거대한 대규모 구조물이다. 이러한 상호 연결된 영역은 거미줄과 같은 패턴을 형성하며 우주에 존재하는 것으로 알려진 가장 큰 구조를 나타낸다. 수억 광년에서 수십억 광년에 이르는 엄청난 거리에 걸쳐 있으며, 이 중 헤라클레스-코로나 보레알리스 만리장성은 우주에서 가장 큰 초은하단 Complex이다. 관측 가능한 우주 내에서 발견되는 은하의 개수는 약 2천억 개이며 그들이 포함된 우주의 크기는 약 930억 광년이다.
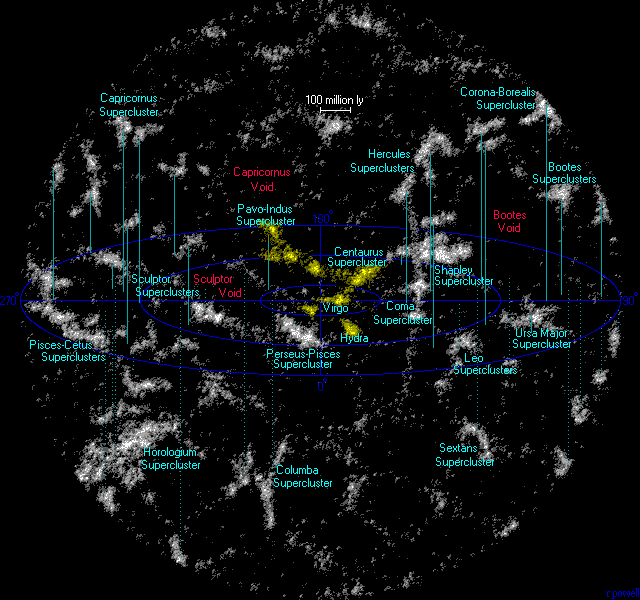
그림 1.5. 가까운 초은하단들(노란색 라니아케아 초은하단)
b. 우주의 창조
우리가 살고 있는 우주는 어떻게 시작되었을까? 이 우주는 시작도 없이 영원 전부터 존재해 온 것일까, 아니면 창조주 하나님에 의해 창조된 것일까? 많은 사람들이 궁금해하는 이 질문에 답하기 위해, 천문학적으로 밝혀진 우주의 기원과 성경 창세기에 기록된 우주의 창조 기록을 살펴보고, 이 둘이 서로 일치하는지 알아보도록 하자.
i. 천문학적으로 밝혀진 우주의 창조
우주의 기원에 대해 가장 널리 지지를 받는 이론은 빅뱅 이론이다. 빅뱅에 의하면, 우리 우주는 약 138억 년 전에 한 특이점의 폭발로부터 시작되었다. 이 사실은 자연스럽게 하나의 흥미로운 질문을 제기한다: 빅뱅 이전에는 무엇이 존재하였을까? 최신 우주론에 의하면 빅뱅 이전에는 우주가 아무것도 존재하지 않는 진공상테였으며 이 진공이 양자 요동(Quantum fluctuation)을 하고 있는 상태에서 어느 한 특이점의 폭발로부터 우주가 시작되었다고 한다.
폴 디랙 이전에는 진공을 아무것도 없는 공간으로 인식하였다. 하지만 디랙이 상대론과 양자역학을 결합하여 디랙 방정식을 풀자, 진공은 무가 아니라 음의 에너지를 가진 입자로 가득 찬 공간임을 발견하였다. 디랙의 이 발견으로 인해 물리학에서는 진공을 디랙의 바다라고 부른다. 디랙의 바다는 인간의 눈에 정적으로 보이지만 결코 정적이지 않다. 오히려 하이젠베르크의 불확정성 원리에 의해 엄청난 요동을 치고 있다. 물질과 반물질이 순간적으로 쌍생성(Pair creation) 되었다가 바로 쌍소멸(Pair annihilation)을 한다. 그 시간이 10-21초가 되어 인간의 눈에는 보이지 않지만, 그것을 볼 수 있는 카메라가 있다면 그야말로 정신없이 요동치고 있는 진공의 바다를 보는 것 같을 것이다. 이것이 바로 ‘양자 요동’이다.
그림 1.6. 양자 요동을 치고있는 진공의 3차원 모델
이렇게 양자 요동이 치고 있는 어느 순간 우주의 한 특이점(Singular point)에서 큰 폭발이 일어나면서 우주가 시작되었다. 이 폭발을 잘 알려진 빅뱅(Big Bang)이라 부른다.
빅뱅 직후 우주는 엄청나게 높은 온도와 밀도로 인하여 아주 짧은 시간 동안 많은 변화를 거친다. 우주가 시작된 직 후 플랑크 시간인 10-43초부터 10-36초까지는 우주를 지배하는 모든 힘들이 통일된 대통일 시대를 거치며, 10-36초부터 10-32초까지는 인플레이션 시대, 10-32초부터 10-12초까지는 전기 약력 시대, 10-12초부터 10-6초 사이에서는 쿼크 시대, 10-6초부터 1초 사이에서는 강입자 시대, 1초부터 10초 사이에는 경입자 시대를 거친다.
경입자 시대 말미에 우주의 온도가 109도 정도로 식으면 아주 중요한 사건이 일어난다. 경입자와 반경입자(주로 전자와 양전자)가 만나 쌍소멸을 하면서 온 우주가 광자(빛 입자)로 충만해지는데 이때를 광자 시대(photon epoch)라 부른다. 광자 시대는 우주의 나이가 38만 년이 될 때까지 지속된다. 광자 시대 초기 약 20초부터 20분 사이에는 핵합성 시대를 거치면서 양성자, 중성자, 헬륨이 생성되어 물질이 만들어질 준비를 한다. 빅뱅 직후부터 광자 시대까지를 복사 지배 시대(radiation-dominated era)라 부르는데 시간상으로 보면 복사 지배 시대의 거의 100%는 광자가 지배하는 광자 시대로 볼 수가 있다. 광자 시대에 생성된 광자는 플라즈마 상태의 자유전자와 양성자에 의해 산란되어 자유롭게 이동하지 못하고 있다. 따라서 우주는 아직 잘 관측되지 않는 불투명 상태로 존재하게 된다.
광자 시대 말미에 우주의 온도가 어느정도 내려가기 시작하면서 재결합 시대가 시작된다. 이때 핵합성 시대에 만들어진 자유 전자와 양성자가 결합하여 중성 수소가 만들어진다. 그렇게 되면 플라즈마로 가득 찬 우주가 서서히 투명해지면서 우리가 하늘이라고 부를 수 있는 공간이 나타난다. 이렇게 되면 광자 시대에 생성되었지만, 플라즈마에 막혔던 광자들이 비로소 투명해진 우주를 활보하며 우리에게 관측되기 시작한다. 이렇게 자유롭게 움직이는 광자는 매우 밝은 빛으로 관측되었으며 현재의 우주 마이크로파 배경 복사의 기원이 된다.
재결합 시대에 물질들이 만들어지면서 물질 지배 시대(matter-dominated era)로 들어가게 되는데 이때 형성된 물질이 현재 우리가 보는 별과 은하들을 생성하는 재료가 되었다. 별과 은하들이 생성된 이후 우주는 빅뱅의 여파로 계속 팽창한다. 우주의 나이가 98억 년이 되었을 때는 눈에 보이지 않는 암흑 에너지가 우주를 지배하는데 이때를 암흑 에너지 지배 시대(dark energy-dominated era)라 부르며 은하들의 거리가 서로 멀어지면서 가속 팽창을 하고 있는 것이 현재 우주의 상태이다.
ii. 우주의 운명 (다시 시작?)
가속 팽창을 하고 있는 우주의 운명은 어떻게 될까? 우리 우주의 운명은 우주의 전체 밀도에 달려 있다. 우주의 밀도가 임계 밀도(10–29g/cm3)보다 큰 경우(닫힌 우주) 우주는 팽창을 지속하다가 어떤 시점에서 팽창을 멈추고 다시 수축하게 된다. 이것을 빅 크런치(Big Crunch)라 부른다. 반대로 밀도가 임계 밀도보다 적은 경우(열린 우주) 우주는 가속 팽창을 계속하여 결국은 빅립(Big Rip)으로 알려진 시나리오로 이어진다. 이 경우 팽창이 진행됨에 따라 우주의 온도는 점차 낮아지고 별 생성에 필요한 성간 매질이 고갈되면서 새로운 별들이 더 이상 생성되지 않는다. 기존에 있던 별들도 연료가 고갈되어 빛을 발하지 않게 된다.
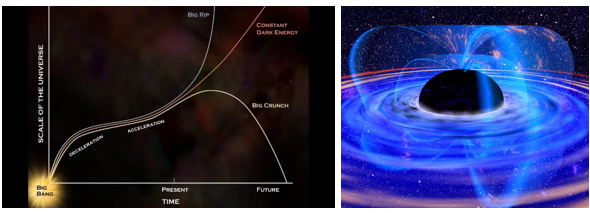
그림 1.7. 우주의 운명과 증발하는 블랙홀
그 후 우주의 나이가 약 1032년이 되면 대통일이론이 예측한 대로 양성자 붕괴가 시작되어 물질이 점차 사라진다. 약 1043년이 지나면 블랙홀마저 호킹 복사를 통해 증발하기 시작한다. 모든 입자가 붕괴하고 블랙홀이 증발되면, 우주는 복사로 가득한 암흑의 시기를 맞이하게 된다. 우주의 온도는 절대 영도로 냉각되고, 결국 모든 것이 어둡고 텅 비어 빅뱅 이전의 양자 요동을 겪고 있는 우주의 상태와 비슷해진다.
우주의 운명에 대한 또 다른 이론은 우주가 순환된다는 순환 우주론이다. 최근 지구에서 북두칠성 방향으로 70억 광년 떨어진 곳에서 두 개의 우주 거대 구조가 발견되었다. 2022년에 발견된 Giant Arc와 2024년에 발견된 Big Ring이다.
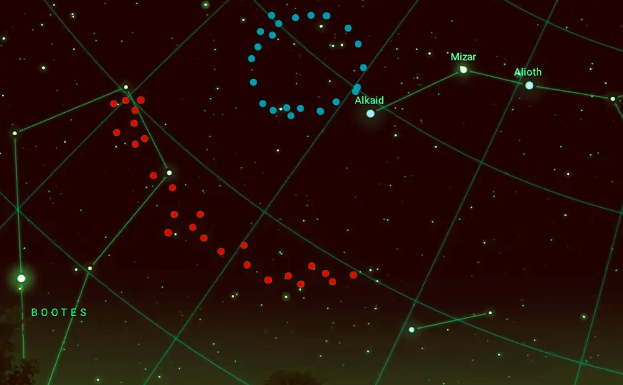
그림 1.8. 빅 링(파란색) 및 빅 아크(빨간색)
이 두 거대 구조는 우주원리(Cosmological Principle)의 균질성을 위배할 수도 있으며 어떤 적절한 설명이 필요하다. 가능한 설명으로는 우주 끈(Cosmic string) 또는 이전 빅뱅에서 생긴 초거대 블랙홀의 잔해가 아닌가 추측한다. 호킹 복사에 의한 블랙홀의 증발속도는 생각보다도 아주 느리다. 태양 질량 정도의 블랙홀 증발 시간은 약 1067년이고 초거대 블랙홀의 증발 시간은 약 10100년이 걸린다. 그런데 이 블랙홀들이 만나 충돌하면 아주 강력한 중력파가 발생한다. 이번에 발견된 빅 링이 이전의 우주에서 발생한 중력파의 흔적(Hawking Point)일 수도 있다는 것이다. 이 해석은 로저 펜로즈의 등각 순환 우주론(CCC)과 관련이 있다. CCC는 일반 상대성 이론에 기반한 우주론적 모델로, 우주가 팽창하며 모든 물질이 소멸하고 블랙홀만 남는 상태가 될 때, 다시 새로운 빅뱅이 시작된다는 가설이다.
개인적으로 CCC가 흥미로운 이유는, 은하 진화의 몇 가지 문제를 해결할 가능성을 제시하기 때문이다. 예를 들어, 블랙홀의 질량과 항성 속도 분산 사이에는 상관관계(M-sigma relation)가 존재하며, 블랙홀의 질량은 일반적으로 은하 질량의 약 0.1% 정도에 해당한다. 그런데 우주 초기의 블랙홀 질량을 보면 이 비율이 맞지 않는다. 예를 들어, 빅뱅 후 약 5억 년이 지난 은하 UHZ1의 블랙홀은 은하 전체 질량보다 크다. 이를 설명하기 위해 여러 가설이 제시되었지만, CCC를 적용하면 쉽게 해결할 수 있다. 즉, 이전 우주에서 생성된 블랙홀이 새로운 우주의 "씨앗 블랙홀"이 되어 주변의 가스와 먼지를 모으고 이를 통해 은하가 형성되었다고 가정하면 초기 우주의 비정상적으로 큰 블랙홀 질량 문제를 효과적으로 설명할 수 있다.
그렇다면, 팽창하는 우주 속에서 다시 새로운 우주가 태어난다는 말은 무슨 의미일까? 3차원 공간에서 어떤 물질들이 흩어진다는 것은 한 점에 모인다는 의미의 반대말이다. 하지만 4차원 공간에서는 바라본 3차원의 흩어짐은 4차원 공간의 위상 구조에 따라 한 특이점으로 수렴하는 구조로 되어 있을지도 모른다. 예를 들어, 클라인 병(Klein bottle)의 내부 표면을 통해 나온 물체가 입구에서는 하나의 점으로 수렴하여 들어가는 것처럼 3차원에서의 팽창이 4차원에서는 수렴하는 형태로 나타날 수도 있다. 만약 그렇게 되면, 등각 순환 우주론은 자연스럽게 설명될 수 있다. 어쩌면 4차원 공간에 사는 초등학생도 4차원에서 바라본 3차원의 흩어짐은 4차원에서 한 특이점에 모일 수 있다는 것을 상식으로 알고 있을지도 모른다.
이 경우, 한없이 팽창하는 우주는 거대한 진공의 바다로 변하며 양자 요동이 일어나면서 다음 우주를 준비한다. 이렇게 팽창하는 우주가 한 특이점에 계속 몰리면 특이점의 진공 에너지 밀도가 점점 올라가고 이렇게 올라가는 진공 에너지 밀도를 특이점에서 다 수용할 수 없어 대폭발이 일어나게 되면 이게 다음 우주의 빅뱅일 수도 있다는 이야기다. 빅뱅 이후에도 특이점으로 계속 진공 에너지가 몰려오면 그게 암흑 에너지의 기원이 될 수가 있고 새로 생긴 우주 팽창의 동력원이 될 수도 있을 것이다.

그림 1.9. 등각순환 우주론
iii. 기독교 성경에 기술된 우주의 창조 이야기
기독교의 경전인 성경은 하나님의 말씀이 기록된 책이며 역사상 가장 많이 읽힌 베스트셀러이다. 성경의 첫 번째 책인 창세기에는 천지 만물의 창조에 대한 기록이 담겨 있다. 이 장에서는 창세기에 기록된 우주 창조가 천문학적 사실과 어떻게 일치하는지 알아보도록 하자.
a) 하나님의 창조 선언
창세기 1장 1절에 다음과 같이 기록되어 있다.
“태초에 하나님이 천지를 창조하시니라”(창세기 1:1).
위 구절에서 볼 수 있듯이 성경은 하나님께서 직접 우주를 창조하셨다고 구체적으로 기록하고 있다. 이어서 우주 창조에 대한 설명이 계속되는데 창세기 1장 2절을 보면 우주가 시작되기 전에는 어떤 상태였는지를 묘사하고 있다.
“땅이 혼돈하고 공허하며 흑암이 깊음 위에 있고 하나님의 영은 수면 위에 운행하시니라”(창세기 1:2).
여기서 "땅"은 하나님께서 나중에 형성하실 물질, 즉 baryonic matter을 의미한다. ‘혼돈하고’는 영어 성경에 ‘without form’으로 번역된다. 따라서 ‘땅이 혼돈하고’라는 표현은 태초에 어떤 물질도 형성되지 않았음을 의미한다. 다음으로, ‘공허하고’는 영어 성경에서 ‘void’로 번역되며 아무것도 존재하지 않는 진공 상태를 의미한다. 즉, 태초에는 아무것도 없는 진공 상태가 존재했음을 나타낸다. 여기까지는 해석에 무리가 없을 것으로 생각한다.
그런데 그다음 문장 ‘흑암이 깊음 위에 있고’가 심오한 의미를 내포하고 있다. ‘흑암’은 히브리어로 ‘וחש’이며, 이는 빛이 창조되기 이전의 근본적 암흑상태를 의미한다. 이어지는 단어인 ‘깊음’이 아주 핵심적인 의미를 갖는다. 깊음의 히브리어는 ‘תהום’인데, 이는 ‘요동치다’ 또는 ‘부글거리다’라는 뜻을 가진 ‘הום’에서 유래한 단어로 깊이를 알 수 없는 바다 또는 공간이 요동치는 상태를 나타낸다.
이것을 앞의 문장들과 연결하면 ‘아무것도 없는 암흑상태의 진공이 요동치고 있음’을 의미하게 된다. 이 해석은 앞서 언급한 천문학적 사실, 즉 빅뱅이 시작되기 전 '암흑 상태의 진공이 양자 요동을 치고 있는 상태'를 정확하게 표현하고 있다.
b) 창조 첫째 날 빛의 창조
진공의 양자 요동을 기록한 창세기 1장 2절에 이어, 창세기 1장 3절에서는 하나님께서 창조 첫째 날에 행하신 놀라운 일이 기록되어 있다.
“하나님이 이르시되 빛이 있으라 하시니 빛이 있었고”(창세기 1:3).
이 구절은 하나님께서 우주를 창조하실 때 빛을 먼저 창조하셨다고 기술하고 있다. 이 창세기에 기록된 빛의 창조는 빅뱅 직후 광자 시대에 빛이 생성되어 온 우주를 채운 천문학적 사실과 정확히 일치한다. 광자 시대에 생성된 빛은 우주가 투명해지고 하늘이 형성되었을 때 우리에게 관측이 되었다.
c) 창조 둘째 날 하늘의 창조
창세기 창조 둘째 날의 주요 사건은 하늘의 창조이다.
"하나님이 궁창을 만드사..., 하나님이 궁창을 하늘이라 부르시니라...."(창세기 1:7, 8).
창세기에 기록된 하늘의 창조는 빅뱅에 의해 발생한 광자 시대 다음에 오는 재결합 시대와 연관이 되어있다. 광자 시대의 우주는 밀도가 높고 뜨거운 플라즈마로 가득 찬 상태였다. 이 플라즈마는 광자를 산란시켜 광자가 자유롭게 이동하는 것을 방해하여 우주를 불투명한 상태로 만들었기에 우리가 하늘이라고 부를 수 있는 공간이 존재하지 않았다. 그러나 재결합 시대에 우주의 온도가 점점 내려가자, 전자와 양성자가 결합하여 중성 수소를 형성하였다. 이렇게 되자 플라즈마에 막혀있던 광자가 자유롭게 이동하게 되고 그 결과 우리가 하늘이라고 부를 수 있는 공간이 나타나게 된다.
따라서 창세기 창조 둘째 날 하늘의 창조는 재결합 시대에 하늘이 나타난 사건을 정확히 묘사하고 있다.
다음 표는 성경에 기록된 우주의 창조와 천문학적으로 밝혀진 우주의 생성 과정을 요약한 것이다. 이 표는 창세기의 창조 기록이 천문학적 사실과 놀라울 정도로 정확히 일치함을 보여주며, 하나님께서 이러한 사실들을 과학이 밝혀내기 훨씬 이전에 이미 성경을 통해 알려주셨음을 확인시켜준다.
성경 창세기 | 천문학 |
진공의 양자 요동 (창조 이전 - 창 1:2) | 진공의 양자 요동 (빅뱅 이전) |
빛의 창조 (창조 첫째 날 - 창 1:3) | 빛의 창조 (광자 시대) |
하늘의 창조 (창조 둘째 날 - 창 1:7-8) | 하늘의 창조 (재결합 시대) |
표 1.1. 성경 창세기와 천문학의 우주 창조 비교
c. 지구와 태양, 어느 것이 먼저 창조되었는가?
창세기에서는 셋째 날에 하나님께서 물과 함께 지구를 창조하시고, 넷째 날에 지구를 비추는 광명체인 태양을 만드셨다고 기록되어 있다. 이 기록은 지구가 태양보다 먼저 창조되었음을 나타내는데, 과연 이것이 천문학적으로 사실인지 알아보도록 하자.
별과 행성은 성간 분자구름(molecular clouds)에서 형성된다. 분자구름의 구성 성분은 약 98%가 가스(수소 약 70%, 헬륨 약 28%)이며 나머지 2%는 먼지(탄소, 질소, 산소, 철 등)로 이루어져 있다. 별과 목성형 행성의 대부분은 가스로 만들어지며, 지구형 행성의 대부분은 먼지로 형성된다.
분자구름 중심에서 중력 수축이 일어나면 원시별(protostar)이 생성된다. 동시에, 원시별 주위에는 행성이 형성되는 요람인 원시 행성계 원반(protoplanetary disk)이 형성된다. 원시별 내부의 온도는 아직 핵융합을 일으킬 정도로 높지 않으며, 중력 수축에 의한 충격파로 온도가 다소 상승하지만, 가시광선으로는 관측되지 않는다. 원시별이 계속해서 서서히 수축하면 전주계열성(pre-main sequence star)으로 진화하고, 하야시 경로(Hayashi track)와 헤니에이 경로(Henyey track)라는 항성 진화 과정을 거친다. 이 단계를 지나 원시별 내부의 온도가 핵융합이 가능한 1천만~2천만 도까지 상승하고 수축이 멈추면 우리가 밤하늘에서 볼 수 있는 광명체, 즉 진정한 별이 탄생한다. 이때부터 별의 생애는 주계열성(main sequence star) 단계에 접어들며, 핵융합이 시작되는 시점을 영점 주계열(zero age main sequence)이라고 부른다. 항성 진화 이론과 태양 지진학(helioseismology) 연구에 따르면 태양은 약 4천만~5천만 년 동안 전주계열성 상태를 거친 후 영점 주계열성에 도달하여 핵융합을 시작하고 스스로 빛을 내는 광명체가 된다.
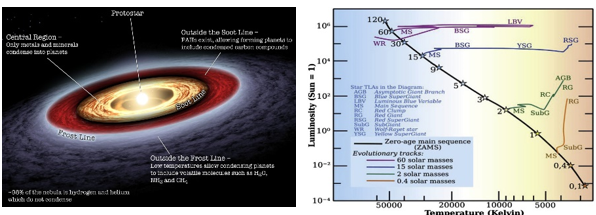
그림 1.10. 원시별과 원시 행성계 원반, 그리고 H-R 다이어그램
행성의 경우는 원시 행성계 원반을 구성하고 있는 가스와 먼지들이 충돌하면서 모래 크기, 자갈 크기, 바위 크기, 수십 또는 수백 킬로미터 크기의 미행성(planetesimal), 그리고 이 미행성들이 뭉친 원시행성(protoplanet)이 만들어지고 행성으로 진화한다. 정리하면, 지구와 태양은 같은 분자구름으로 시작하였지만, 지구는 원시 행성계 원반에서 원시행성을 거쳐 탄생하였고 태양은 전주계열성을 거쳐 주계열성으로 탄생한다.
원시 행성계 원반의 먼지구름에서 행성이 생성되는 과정이 최근에야 활발히 연구가 진행되고 있다. 최근 발표된 논문들을 보면 원시 행성계 원반의 먼지구름에서 지구형 행성인 수성, 금성, 지구, 화성의 질량을 합친 질량의 자갈들이 중심의 전주계열성으로부터 4천문단위 내에 형성되는데 걸리는 기간은 약 30만 년이다. 또한, 1밀리미터 크기의 모래들로부터 원시행성을 거쳐 지구 크기의 행성이 탄생하는 기간은 약 수백만 년이 걸린다고 예측하였다.
이러한 행성 생성 이론들은 관측을 통해 확인될 수 있다. 다음 이미지는 세계 최대 서브밀리미터 망원경인 ALMA로 관측한 황소자리 HL Tau(좌)와 센타우루스자리 PDS 70(우)이다. HL Tau의 질량은 태양의 약 2배 정도이고 나이는 백만 년 정도이며 중심에는 전주계열성 별이 형성되고 있다. 그런데 이렇게 별이 형성되는 초기 단계에 이미 여러 행성이 만들어져 그 주위를 돌고 있는 것을 보여 준다. 전주계열성 주위의 원반을 보면 여러 개의 갭(gap)들이 보이는데 이 겝들이 바로 행성이 형성되어 주위의 먼지구름들을 휩쓸고 지나간 흔적이다.
PDS 70의 질량은 태양의 0.76배 정도이며 나이는 540만 년 정도이고 중심에는 전주계열성이 성장하고 있다. PDS 70의 안쪽 궤도를 보면 2개의 목성형 행성 PDS 70b와 PDS 70c가 형성되어 공전하고 있는 것이 관측되었으며 HL Tau에서 보였던 먼지구름들이 이 2개의 행성에 의해 거의 다 제거되었음을 보여 준다. 2023년 제임스웹 우주망원경으로 중심 부분을 분광 관측하자 그림의 스펙트럼에 나타난 것처럼 다량의 물이 관측되었으며 관측된 영역은 지구형 행성들이 생성되는 장소로 그 속에 2개 이상의 지구형 행성들이 형성되어 있음을 시사하였다.
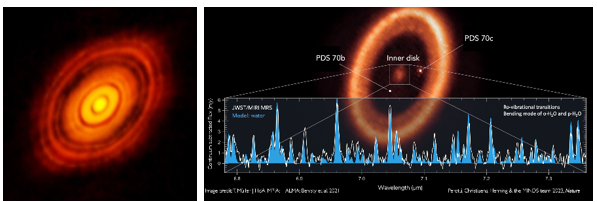
그림 1.11. HL 타우와 PDS 70
여기서 중요한 사실은 원시 행성계 원반의 먼지구름에서 약 540만 년이 지나자, 물과 함께 지구형 행성들이 생성되었다는 점인데, 이는 창세기 창조 셋째 날에 물과 지구가 함께 창조되었다는 기록과 일치한다. 이때까지 걸린 시간이 540만 년이지만 길게 잡아 천만년이 걸렸다 하더라도 이 기간은 태양이 별이 되는 기간인 4천만~5천만 년보다 훨씬 짧다는 것을 보여 준다.
따라서 창세기에 기록된 지구가 먼저 창조되었고 그다음 태양이 창조되었다는 기록이 천문관측을 통해 사실임을 입증해 준다.
하나님께서 셋째 날에 행하신 또 다른 주요 사건은 식물과 나무의 창조다. 무신론자와 진화론자들은 종종 "지구가 먼저 창조되고 그 후에 태양이 창조되었다면 지구와 함께 창조된 식물들은 태양이 없는데 어떻게 살아남았는가?"라는 의문을 제기한다. 그러나 이 문제는 별의 진화 과정을 알면 쉽게 해결된다. 중력 수축으로 별이 만들어지고 있는 T Tauri 별의 표면 온도는 약 4,000~5,000도 정도가 되며 흑체복사 피크가 가시광선 영역에 위치한다. 또한 전주계열성인 T Tauri 별 단계에 있는 태양의 크기가 현재 주계열성별인 태양보다 몇 배 정도 커서 지구상의 식물들이 광합성을 하는 데 충분한 에너지를 공급해 줄 수가 있었다. 따라서 태양이 주계열성에 도달하기 전에도 식물의 생존에는 아무런 문제가 없었다.
d. 지구의 나이는 6,000년인가?
젊은 지구 창조론자들은 성경 창세기의 창조 기록을 피조물의 입장에서 해석하여 지구와 우주의 나이가 6,000년 정도라고 주장한다. 하지만 지질학, 천문학, 물리학 등 다양한 분야의 여러 과학적 증거에 따르면 지구의 나이는 약 46억 년, 우주의 나이는 약 138억이 된다. 이러한 과학적 사실에도 불구하고 젊은 지구 창조론자들은 여전히 지구의 나이가 6,000년이라고 주장한다. 본론으로 들어가기 전에, 먼저 지구와 우주의 나이가 적어도 수백만 년이라는 사실을 쉽게 이해할 수 있는 예를 하나씩 살펴보도록 하자.
지구의 나이가 최소 수백만 년이 되었다는 것은 판구조론 인정하면 된다. 지구의 지각은 여러개의 판으로 이루어져 있으며 이 판들의 움직임에 의해 지진이 일어난다. 이 사실을 부인하는 사람은 없을 것이다. 열점(hot spot)은 지각 아래에 있는 맨틀 심부에서 마그마가 흘러나오는 지점으로, 그 중심부가 고정되어 있다. 마그마가 지각 위로 분출되어 식으면 땅이 만들어 지는데 그 대표적인 예가 하와이 제도이다. 하와이 본섬인 빅아일랜드의 킬라우에아에는 지금도 화산활동이 활발하게 일어나고 있으며 분출된 마그마가 바닷물에 식으면서 땅이 만들어진다. 이렇게 형성된 땅은 판구조론에 의해 북서쪽으로 연간 약 7~10cm씩 이동하며, 이 과정이 반복되면서 하와이 제도의 여러 섬이 형성되었다. 이는 현재도 관측할 수 있는 부정할 수 없는 사실이다.
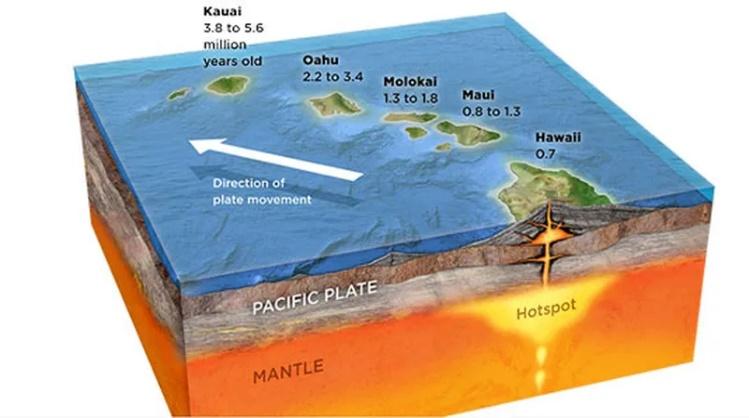
그림 1.12. 화산활동으로 형성된 하와이 제도
지각판의 이동 속도를 고려한 하와이 섬들의 나이는 북서쪽 방향으로 빅아일랜드는 40만 년, 마우이는 1백만 년, 몰로카이는 1.5~2백만 년, 와이키키가 있는 오아후는 3~4백만 년, 카우아이는 5백만 년쯤 되었다. 빅 아일랜드의 경우 땅의 상당 부분이 여전히 검은 화산토로 덮여 있어 풍화가 거의 일어나지 않았음을 알 수가 있다. 반면 카우아이는 상대적으로 풍화가 많이 진행되어 수목들이 잘 자라고 있어 정원의 섬으로 불린다. 이 사실들은 지구의 나이가 최소한 몇백만 년은 되었다는 것을 직접적으로 증거하고 있다.
우주의 나이가 최소 수백만 년 이상이라는 것은 빛이 1초에 30만km를 이동한다는 것을 인정하면 된다. 이 사실 역시 부정할 수 없다. 태양은 지구로부터 1억 5천만km 떨어져 있다. 우리가 지금 따뜻한 햇빛을 받고 있다는 것은 그 빛이 태양에서 8.3분 전에 생성된 빛이라는 것을 의미한다. 태양은 달보다 약 400배 정도 큰데 멀리 떨어져 있어 시직경이 달과 비슷하게 보인다. 안드로메다은하는 우리은하와 크기가 비슷하지만 250만 광년 떨어져 있어 달의 4배 크기 정도로 적게 보인다. 우리가 안드로메다은하를 볼 수 있다는 것은 우리에게 도달한 빛이 250만 년 전에 안드로메다에서 생성되어 우리에게 도달하였다는 것을 의미한다. 안드로메다은하를 보았다면 이 사실을 부정하지 못할 것이다. 이 사실은 우주가 최소한 수백만 년은 되었다는 것을 직접적으로 시사하는 증거이다.
이러한 사실에도 불구하고 여전히 지구의 나이가 6,000년이라고 주장한다면 복음을 전파하는 데 도움이 되기보다는 걸림돌이 될 수 있으며, 잠재적으로 많은 사람들을 복음으로부터 멀어지게 할 수 있다. 따라서 무리하게 젊은 지구 창조론을 주장하기보다는 성경 창세기를 주의 깊게 읽고 해결책을 찾는 것이 더 이성적일 수 있을 것이다.
인간에게 시간은 변할 수 없는 물리량이며 항상 현재에서 미래로 흐르는 것으로 인식되어 왔다. , 시간에 대한 이런 고정 관념을 제거하고 이 문제에 접근해 보면 아마 어떤 해법이 생길 수도 있을 것이다. 이 사실을 염두에 두고 이 문제에 대해 좀 더 자세히 알아보도록 하자.
i. 창세기의 창조 6일
먼저 창세기의 기록을 바탕으로 우주의 나이를 추정해 보도록 하자. 창세기에 따르면 하나님은 6일에 걸쳐 우주와 그 안의 모든 것을 창조하셨다. 하나님께서 처음 창조하신 인간인 아담은 몇 살까지 살았고 그 자손들은 몇 살까지 살았는지 그 족보가 성경의 창세기 5장에 자세히 기록되어 있다.
“이것은 아담의 계보를 적은 책이니라…노아가 오백 세 된 후에 셈과 함과 야벳을 낳았더라”(창세기 5:1-32).
아담으로부터 시작하여 그의 자손인 노아 시대 홍수 때까지의 기간을 성경에 기록된 대로 합산하면 1,656년이 된다. 노아 홍수의 시기에 대해서는 다양한 의견들이 있지만 여러 자료를 참고하면 노아 홍수는 지금으로부터 약 2,400년 전에 발생한 것으로 추정된다. 따라서 아담이 창조된 후 노아 홍수 때까지가 1,656년이고 노아 홍수 때부터 지금까지 약 4,400년이 지났으므로 성경에 언급된 우주의 나이는 창조 7일 +1,656년 +4,400년으로 약 6,056년이 된다. 이것이 바로 지구의 나이가 6,000년이라고 주장하는 젊은 지구 창조론자들의 근거가 된다.
이 문제를 해결하기 위해 다시 한번 창세기를 주의 깊게 살펴보도록 하자. 아담 이후의 기록은 정확한 것으로 추정되며, 노아의 홍수가 일어난 시기가 지금으로부터 4,400년 전이든 44,000년 전이든 우리 우주의 나이 138억 년에는 어떤 영향도 미치지 않는다. 그렇다면 창세기에 나타난 우주의 나이가 틀렸을까 아니면 창세기 기록 안에 이 문제를 풀 수 있는 열쇠가 들어 있을까? 아마 창세기 1장을 주의 깊게 읽고 약간의 천문학적 지식이 있는 사람은 창세기의 우주 창조 최초 7일에 그 열쇠가 들어 있음을 추측할 수 있을 것이다.
그 추론은 다음과 같다. 피조물의 입장에서 정의하는 하루는 피조물이 사는 행성 자전 속도에 의해 정의된다. 인간이 금성에 창조되었다면 하루는 243일이고 목성에 창조되었으면 하루는 10시간이다. 인간이 지구에서 창조되었기에 우리의 하루는 지구가 한 번 자전하는 데 걸리는 시간인 24시간이다. 또한 인간의 관점에서 하루를 정의하기 위해서는 먼저 지구가 있어야 하고 지구를 비추는 태양이 있어야 한다. 그런데 지구는 창조 셋째 날에 그리고 태양은 창조 넷째 날에 창조되었다. 하지만, 인간의 하루가 정의되기 이전에 하나님께서는 하루라는 단어를 사용하셨다. 따라서 창세기 창조 7일 동안의 하루는 우리 인간이 정의하는 하루가 아니라 하나님의 관점에서 정의하신 하루였다고 볼 수 있다. 젊은 지구 창조론자들이 창세기의 하루를 인간의 하루로 간주하여 지구의 나이를 6천 년이라고 주장한 오류가 여기서 발생하였으며 지구 나이의 비밀은 바로 이 창조 7일의 시간에 담겨 있다.
그림 1.13. 하루를 정의하기 위해 지구와 태양이 먼저 존재해야 한다
그렇다면 창세기 창조 7일 동안의 시간은 인간의 시간으로는 얼마나 될까? 정확한 답은 알 수 없지만 창세기에 기술된 창조 사건과 빅뱅의 사건을 비교하면 대략적인 시간을 추정할 수 있을 것이다.
창세기 창조 첫째 날의 대표적인 사건은 빛의 창조다. 천문학적으로는 빅뱅 직후 광자 시대가 이 사건에 해당함으로 창조 첫째 날의 시간은 38만 년이 된다. 창조 둘째 날의 대표적인 사건은 하늘의 창조다. 광자 시대 이후의 재결합 시대가 이 사건에 해당함으로 창조 둘째 날의 시간은 10만 년이다. 창조 셋째 날의 대표적인 사건은 지구의 창조다. 앞에서 살펴본대로 지구형 행성이 생성되는 시간이 약 1천만 년이 걸리니까 창조 셋째 날의 시간은 약 1천만 년 보다는 클 것이다. 창조 넷째 날의 대표적인 사건은 태양의 창조다. 태양이 형성되는 시간이 4-5천만 년이 걸리니까 창조 넷째 날의 시간은 4천만 년 이상일 것이다. 이 결과들을 요약하면 다음 표와 같다.
창조 | 창세기 사건 | 천문학 사건 | 시간 |
첫째 날 | 빛의 창조 | 광자 시대 빛의 창조 | 380,000년 |
둘째 날 | 하늘의 창조 | 재결합 시대 하늘의 창조 | 100,000년 |
세쨋 날 | 지구의 창조 | 지구의 창조 | > 1,000만 년 |
넷째 날 | 태양의 창조 | 태양의 창조 | > 4,000만 년 |
표 1.2. 창세기 창조 시간
이렇듯 창조 넷째 날까지만 살펴보더라도 우리는 한 가지 경이로운 사실을 발견하게 된다. 하나님의 관점에서 본 창조의 하루는 인간이 정의한 24시간에 비해 아주 긴 시간이며 또 그 시간이 수십만 년으로 짧기도 하고 4천만 년 보다 더 길기도 하는 등 변한다는 것이다. 이것을 어떻게 이해할 수 있을까? 아마 어떤 관점에서는 이것이 경이로운 결과가 아니라 이미 예상된 결과라고 볼 수가 있다.
ii. 시간의 창조자
창세기에 사용된 '날'의 히브리어는 욤(יום)이다. 욤은 몇 가지 의미로 해석되기도 하는데 그중 기간이 정해지지 않은 시간을 나타내는 의미도 있다. 창조주 하나님께 있어 시간은, 시간의 한계 속에 살아가는 인간과는 전혀 다른 개념이다. 왜냐하면, 하나님께서는 시간을 창조하시고 다스리시기 때문이다. 조금 생소하게 들릴 수 있겠지만, 성경에는 하나님께서 시간을 다스리는 사건이 여러 차례 등장한다.
먼저 신약 성경에 있는 베드로후서를 보면 다음과 같은 성경 구절이 나온다.
“사랑하는 자들아, 주께는 하루가 천 년 같고 천 년이 하루 같다는 이 한 가지를 잊지 말라.”(베드로후서 3:8)
이 구절은 하나님의 약속을 기다리는 자들이 조급해지지 말고 인내로 기다릴 것을 권면하는 의미도 있지만, 하나님의 시간 개념은 인간의 시간 개념과는 전혀 다르며 하나님께서는 시간을 늘리거나 줄일 수도 있으실 분임을 시사하는 성구라고 생각한다.
우리는 시간이 고정된 물리양이 아니라는 것을 알고 있다. 특수 상대성 이론에 따르면, 움직이는 관찰자의 시간은 동일한 관성 프레임에서 정지한 관찰자보다 더 느리게 흐른다(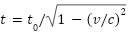 ). 또한, 일반 상대성 이론에서는 강한 중력장하에서 시간이 더 느리게 흐른다는 것을 알고있다(
). 또한, 일반 상대성 이론에서는 강한 중력장하에서 시간이 더 느리게 흐른다는 것을 알고있다(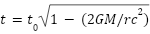 ). 이렇듯 시간의 늘임은 상대론으로 설명이 가능하지만 시간의 줄임은 현재 인간의 과학 지식으로는 이해하기 어려운 부분이다.
). 이렇듯 시간의 늘임은 상대론으로 설명이 가능하지만 시간의 줄임은 현재 인간의 과학 지식으로는 이해하기 어려운 부분이다.

그림 1.14. 시간 팽창을 나타내는 그림
그런데 성경에는 하나님께서 시간을 자유롭게 늘리거나 줄이실 뿐만 아니라, 심지어 시간을 멈추게 하실 수도 있음을 시사하는 사건이 기록되어 있다. 구약의 여호수아서를 보면 다음과 같은 구절이 나온다.
“태양이 중천에 머물러서 거의 종일토록 속히 내려가지 아니하였다”(여호수아 10:13).
이 구절은 여호수아가 아모리 사람들과 전쟁하던 중, 그의 기도에 대한 하나님의 응답으로 태양이 중천에 머물러 거의 하루 동안 지지 않았다는 사건을 기록하고 있다.
그러나 이보다 더욱 놀라운 사건이 구약 열왕기하에 등장한다. 그것은 하나님께서 중병으로 죽을 위기에 처한 히스기야 왕의 간절한 기도를 들으시고, 그에게 15년의 생명을 더 허락하신 표징으로 보여주신 시간의 역전 현상이다.
“선지자 이사야가 여호와께 간구하매 아하스의 해시계 위에 나아갔던 해 그림자를 십도 뒤로 물러가게 하셨더라”(열왕기하 20:11).
여기서 언급한 해시계는 아하스 왕 때 만들어진 것으로 하나님께서 이 해시계의 시간을 열 계단 과거로 되돌리셨다는 말이다. 이 시간의 역전 현상은 시간의 정지나 줄임보다 더 이해하기가 힘든 사실이다. 하지만 시간을 만드시고 그 시간을 빠르게, 느리게, 또는 정지하게 하실 수 있는 하나님께서 시간을 거꾸로 흐르게 하시는 것 역시, 마치 게임기의 조이스틱을 반대로 돌리시는 것처럼 손쉽게 하실 수 있는 일일 것이다.

그림 1.15. 아하스의 해시계 모델
e. 미세 조정된 우주
오케스트라 연주회에 가면, 연주가 시작되기 전에 모든 단원이 자신이 연주할 악기를 조율하는 모습을 보게 된다. 이때, 오보에의 ‘라(A)’ 음을 기준으로 모든 악기가 조율된다. 이는 오보에가 다른 악기들보다 고음에서도 균형 잡힌 음색을 가지며, 상대적으로 안정적인 소리를 내기 때문이다. 또한 ‘라(A)’ 음을 기준으로 삼는 것은 국제적으로 표준화된 조율 기준이 440Hz로 설정되어 있으며, 이 음이 사람의 귀에 가장 잘 들리는 음색이기 때문이라고 한다. 이렇게 조율이 끝난 후, 지휘자가 등장하여 지정된 곡을 연주한다. 만약 악기들이 조율되지 않은 상태에서 연주가 시작된다면 어떻게 될까? 아마도 말 그대로 불협화음의 연주회가 될 것이다.
미세 조정된 우주(Fine-tuned universe)란 단어의 의미 그대로 우주를 구성하고 운행하는 여러 물리 상수들이 우주의 유지와 생명체의 생존을 위해 극도로 정밀하게 조율되어 있다는 사실을 의미한다.
앞서 언급했듯이, 빅뱅으로 우주가 생성될 때 우주의 질량 밀도가 임계 질량밀도보다 컸다면 우주는 생성되자마자 수축했을 것이고, 반대로 임계 질량밀도보다 작았다면 급속도로 팽창하여 별이나 은하, 그리고 생명체가 탄생할 수 없었을 것이다. 그러나 우리 우주의 질량 밀도는 임계 질량밀도와 거의 일치하도록 절묘하게 조정되어, 현재의 우주가 형성되고 유지될 수 있었다.
펜로즈는 그의 책 《The Emperor’s New Mind》(pp 440-447)에서 블랙홀 엔트로피에 대한 버켄슈타인-호킹 공식을 사용하여 빅뱅 직후 우주의 질량밀도가 임계 질량밀도와 비슷할 확률을 계산하였는데 그 확률은 10의 10123 지수승분의 1이었다. 이 확률은 전 세계 바닷가에 있는 모래 중에서 어떤 특정한 모래 하나를 선택하는 것보다 훨씬 더 희박한 확률이며 결코 자연적으로 일어날 수 없는 확률이다.
그런데, 그렇게 탄생한 우주가 아무 이상 없이 잘 운행되고 있는 배후에는 또 다른 주인공들이 있다. 그들은 바로 물리학의 근본적인 상수들이며 그 값들도 생명체의 존재를 위해 절묘하게 설정되어 있다. 우주를 떠받들고 있는 대표적인 물리 상수들은 중력 상수, 빛의 속도, 플랑크 상수, 볼츠만 상수, 유전 상수(vacuum permittivity), 미세구조 상수(fine-structure constant) 등이 있다. 그런데 이 상수들의 값이 지금의 값과 조금만 달랐다면 우리는 이 세상에 존재하지 못하였을 것이다.
가령 중력 상수가 지금보다 적었다면 별과 은하들의 형성이 불가능하였을 것이고 지금보다 컸다면 우리가 살 수 있는 지구 같은 행성들이 생겨나기 힘들었을 것이다. 플랑크 상수의 경우 지금보다 컸더라면 지구에 도달하는 태양의 에너지가 적었을 것이고 이로 말미암아 심각한 기후 변화가 왔을 것이다. 또한 물질들의 결합력이 약해져 쉽게 파괴되고 광합성 효율이 저하되어 식물들의 생존에 영향을 미쳤을 것이다.
그런데 이러한 물리 상수 중에서 물리학자들의 관심을 가장 많이 끈 상수가 다음에 표시된 미세구조 상수 알파이다.
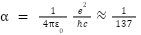
이 상수는 약방의 감초처럼 양자역학의 여러 곳에서 자주 나오며 다른 상수들과 달리 차원이 없는 상수이고 그 값은 약 1/137이 된다. 이 상수의 물리학적 의미는 전자와 양성자 또는 두 전자 간의 전자기력 세기를 나타낸다.
그런데, 이 상수의 값이 1/137보다 컸다면 원자들 간의 상호작용이 강하게 일어나 전자가 원자핵에 더 단단히 결합되고 원자의 크기는 줄어들며 중원소들의 형성이 용이한 반면 수소 같은 원소는 가벼운 원소들은 생성되기 어렵다. 수소는 거의 모든 별의 구성요소이고 핵융합의 원료이기 때문에 수소의 형성이 불가능하면 별 뿐만 아니라 그 별을 에너지원으로 하는 생명체의 생존 역시 불가능하였을 것이다. 반대로 상수의 값이 1/137보다 적었다면 화학반응이 안정적이지 않고 그나마 화학반응을 통해 만들어진 분자와 물질들이 불안정하여 쉽게 붕괴되는 현상이 일어날 것이다. 이렇게 쉽게 붕괴되는 분자들로는 DNA와 단백질로 구성된 생명체가 만들어질 수 없다.
미세구조 상수가 왜 이런 특정한 값(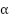 ≈ 1/137)을 가지고 있어 우주 만물을 잘 구성하고 있는지 많은 물리학자를 궁금하게 만들었는데 디랙은 “물리학에서 가장 근본적인 미해결 문제”라고 말하며, 파인먼은 “미세구조 상수는 인간이 이해하지 못한 채로 우리에게 다가오는 마법의 숫자로 물리학의 가장 큰 미스터리다. 당신은 ‘신의 손’이 그 숫자를 썼다고 말할 수 있지만, 우리는 그분이 어떻게 펜을 눌렀는지는 모른다.”라고 말하였다.
≈ 1/137)을 가지고 있어 우주 만물을 잘 구성하고 있는지 많은 물리학자를 궁금하게 만들었는데 디랙은 “물리학에서 가장 근본적인 미해결 문제”라고 말하며, 파인먼은 “미세구조 상수는 인간이 이해하지 못한 채로 우리에게 다가오는 마법의 숫자로 물리학의 가장 큰 미스터리다. 당신은 ‘신의 손’이 그 숫자를 썼다고 말할 수 있지만, 우리는 그분이 어떻게 펜을 눌렀는지는 모른다.”라고 말하였다.
미세구조 상수식을 달리 쓰면 전자의 속도와 빛의 속도비가 된다. 빛이 전자보다 137배 정도 빠르다는 것이다. 또한 미세구조 상수가 관여하는 전자기력은 중력에 비해 약 1036배가 크며 강력에 비해 137배가 적다. 아마, 창조주께서 우주를 만드실 때 물리학 상수들을 개별적으로 설정하지 않으시고 가장 중요한 상수를 먼저 설정하신 후 다른 모든 물리 상수들이 그에 비례하여 자동적으로 설정될 수 있게 하셨는지도 모른다. 그리고 가장 중요하게 설정한 그 상수가 바로 미세구조 상수이며 그 값을 1/137로 설정하셨을 수도 있었을 것이다.
3장의 ‘입자 물리학과 창조론’에서 언급했듯이, 우주의 모든 물질(Baryon)은 17개의 기본 입자들—쿼크, 렙톤, 게이지 보손, 힉스 보손—로 이루어져 있다. 이 기본 입자들은 각기 고유한 질량, 전하, 스핀을 가지고 있는데, 만일 이 값들이 조금이라도 달랐다면 우리가 아는 원자·분자·생명·우주 구조 자체는 존재할 수 없었을 것이다.
예를 들어, 업쿼크와 다운쿼크의 질량 차이가 달라지면 양성자와 중성자의 안정성이 무너져 수소와 원자가 형성되지 못한다. 전자의 질량이 달라지면 원자의 크기와 화학 결합이 불가능해 안정된 분자가 만들어질 수 없으며, 힉스 보손의 질량이 변하면 모든 입자의 질량 분포가 바뀌어 우주의 구조 자체가 달라진다. 또한 양성자와 전자의 전하가 달라지면 전기적으로 중화된 원자가 존재할 수 없고, 쿼크의 전하가 달라지면 양성자와 중성자의 성질이 변해 원자핵 구조가 불가능하다. 더 나아가 전자가 스핀 1/2이 아니라면 파울리 배타 원리가 성립하지 않아 원자 구조가 유지될 수 없고, 보손의 스핀이 달라지면 전기력·강력·약력과 같은 힘의 전달 체계가 무너진다. 마지막으로 힉스 보손이 스핀 0이 아니라면 질량 부여 메커니즘이 작동하지 않아 입자들은 지금처럼 형성될 수 없다.
이처럼 우주의 시작 순간부터 질량 밀도, 기본 물리 상수들, 그리고 17개의 기본 입자들의 고유한 특성에 이르기까지 모든 것이 정밀하게 조율되어 있음을 확인할 수 있다. 임계 질량밀도의 절묘한 일치, 중력·플랑크·미세구조 상수 등 물리 상수들의 섬세한 값, 그리고 쿼크·전자·보손들의 질량과 전하, 스핀의 적절한 배치는 모두 생명과 우주의 존재를 가능하게 하는 필수 조건들이다. 만약 이들 중 단 하나라도 지금과 달랐다면 원자와 분자가 안정적으로 존재할 수 없고, 별과 은하가 형성될 수 없으며, 따라서 생명체 역시 존재할 수 없었을 것이다. 이는 마치 연주회에서 모든 악기가 오보에의 ‘라(A)’ 음에 맞추어 완벽한 조화를 이루듯, 창조주의 정밀한 설계와 의도 아래 모든 물리 상수와 입자들이 미세하게 조율되어 있기 때문에 오늘의 우주가 유지되고 우리가 존재할 수 있음을 보여준다.
이처럼 창조주의 작품들이 어떻게 생겨났고 또 어떤 원리로 작동하는지—중력, 상대성이론, 불확정성 원리, 파울리 배타 원리, 힉스 메커니즘 등—그 일부만 발견해도 세기의 천재로 불리거나 노벨상을 받는다. 그렇다면 이 모든 것을 처음부터 설계하시고 완성하신 창조주 하나님은 얼마나 위대하신가?
2. 하나님의 걸작품 지구
우리가 살고 있는 지구는 인간의 생존에 필요한 물, 공기, 적절한 기후 등 생존에 필수적인 모든 요소를 제공하고 있다. 그러나 같은 지구형 행성인 금성이나 화성의 경우, 물과 공기가 거의 없을 뿐만 아니라 기온이 지나치게 높거나 낮아 생명체가 살아가기 어렵다. 하지만 더 깊이 살펴보면 지구는 단순히 생존에 필수적인 조건을 갖춘 것뿐만 아니라 생명의 존재와 지속적인 보존을 위해 다양한 측면에서 극도로 정밀하게 조정된 특별한 행성임을 알 수 있다. 이러한 조건들은 매우 정교하게 조율되어 있어 마치 미세 조정 우주의 연장선으로 볼 수 있다. 이 장에서는 지구가 우주에서 아주 특별한 행성인 점 10가지에 대해 알아보고자 한다.
a. 태양으로부터 적당한 거리
우리 인간을 비롯한 지구상의 모든 동식물은 물이 없으면 생존할 수가 없다. 그러나 이렇게 생명에 필수적인 물도 행성의 온도가 낮아 얼은 상태로 존재하거나 또는 온도가 너무 높아 뜨겁게 끓는 상태로 존재한다면 생명체의 생존에 부적합하다. 어떤 행성이 그 중심에 있는 별로부터 너무 가까워 물이 끓거나 또는 너무 멀어 물이 어는 곳에 있지 않고 적당한 거리에 위치하여 액체 상태의 물을 가질 수 있는 영역을 생명체 거주가능 영역(habitable zone)이라고 부른다. 태양계 내에서의 생명체 거주가능 영역를 계산해 보면 그 거리는 0.95-1.15천문단위 사이가 된다. 1천문단위는 지구에서 태양까지의 거리인데 공교롭게도 우리 지구는 이 생명체 거주가능 영역의 정중앙에 위치하고 있다. 따라서 지구가 현재의 위치보다 태양 쪽으로 5%만 더 가까웠어도 모든 물이 끓어올랐고 15% 정도만 더 멀었어도 물이 얼어 지구는 생명체가 존재할 수 없는 행성이 되었을 것이다.
다음 그림은 태양계 내의 생명체 거주가능 영역을 표시한 것인데 지구 공전 궤도 주위에 녹색으로 표시된 부분이 생명체 거주가능 영역이다. 태양계 맨 바깥쪽에서 공전하고 있는 해왕성까지의 황도면상에 이 생명체 거주가능 영역이 차지하는 면적의 비율은 단지 0.05%밖에 되지 않는다. 하지만 그림에 표시된 생명체 거주가능 영역은 단지 물의 끓고 얼지 않는 영역을 표시할 뿐이며 실제로 인간과 같은 고등 생명체가 살기에 적합한 온도 범위는 약 5℃에서 50℃ 사이이다. 왜냐하면 너무 춥거나 더울 경우 심각한 기상 재해로 인해 우리의 생존이 위협을 받으며 궁극적으로는 생존에 필수적인 농작물의 재배가 불가능하기 때문이다.
그런데 한 가지 중요한 사실은 지구가 생명체 거주가능 영역 내에 위치해 있더라도 지구의 궤도가 태양을 중심으로 원궤도(그림의 실선)가 아닌 타원궤도(그림의 점선)를 공전하고 있었다면 지구상의 생명체는 생존할 수 없었을 것이다. 왜냐하면 지구가 태양에 가까운 B와 D 지점에서는 지구상의 모든 바닷물이 끓을 정도로 덥고(이심률 > 0.5) 태양에서 먼 A와 C 지점에서는 모든 것이 얼 정도로 추운(이심률 > 0.3) 얼음 행성이 되기 때문이다. 그런데 다행스럽게도 지구는 이심률은 0.017인 거의 원에 가까운 궤도로 공전하고 있기 때문에 이러한 극단적인 기후 변화 없이 생명체가 안정적으로 존재할 수 있는 환경이 유지되고 있다.
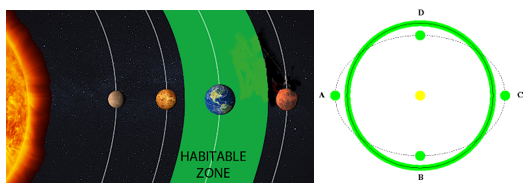
그림 2.1. 태양계의 생명체 거주가능 영역(녹색)
b. 자전축이 적당히 기울어져 있는 지구
지구 자전축의 기울기는 생명체의 생존에 아주 중요한 영향을 미친다. 현재 지구의 자전축이 23.5도로 적당히 기울어져 있기 때문에 우리는 온화한 기후와 사계절을 가질 수 있게 되었다. 그런데 만일 지구의 자전축이 똑바로 서 있거나 아니면 완전히 기울어져 있다면 어떤 일이 벌어질까? 그럴 경우에는 심각한 기상 재해가 발생하여 생명체의 생존을 직접적으로 위협하게 된다.
먼저, 지구의 자전축이 전혀 기울어지지 않고 똑바로 서 있으면 어떤 현상이 일어날지 자세히 알아보자. 그럴 경우 지구상의 어떤 지점이든지 일 년 사시사철 항상 같은 세기의 태양빛을 받게 되며 오직 한 계절만 가질 수 있다. 예를 들어 다음 좌측 그림에 나타난 것처럼 적도지방에 사는 사람들은 지구가 태양 공전 궤도상의 어느 지점에 위치해 있더라도 태양이 항상 중천에서 쨍쨍 내리쪼여 지금보다도 훨씬 더 무덥고 뜨거운 여름만 가지게 될 것이며 반대로 지구의 극지방에 사는 사람들은 지금보다도 훨씬 추운 겨울만 계속되는 기후를 가지게 될 것이다. 이렇게 되면 지구상에서 인간과 생물들이 쾌적하게 거주할 수 있는 지역은 중위도 지역이 되기 때문에 주거 공간이 줄어들고 그에 따른 환경문제가 심각하게 대두될 것이다.
하지만 이보다 훨씬 더 심각한 문제는 기후 변화에 따른 기상 이변 현상이다. 예를 들어, 태풍은 보통 해수면 온도가 약 27도 이상일 때 발생하며, 주로 적도 지방의 해수면 온도가 높은 7~8월에 자주 형성된다. 태풍은 이동하면서 해수면에서 에너지를 흡수해 세력을 키우는데, 만약 태풍이 지나가는 해역의 수온이 낮으면 점차 약화된다. 그러나 엘니뇨 현상 등으로 인해 해수면 온도가 높아지면 태풍은 더욱 강력해질 수 있다. 그런데 자전축이 똑바로 서 있는 경우, 적도 지방의 해수면 온도는 연중 높은 상태를 유지하게 된다. 그렇게 되면 태풍이 일 년 내내 발생할 가능성이 높아지고 더욱 강력한 태풍이 잦아질 것이다. 또한, 해수면 온도가 높으면 해수의 증발이 증가하면서 태풍과 함께 대규모 홍수 피해도 동반될 것이다. 우리나라의 경우, 여름철 한두 달 동안 발생하는 태풍 피해만으로도 큰 영향을 받는데, 만약 일 년 내내 더욱 강력한 태풍과 홍수가 지속된다면 피해 규모는 상상하기 어려울 정도로 커질 것이다. 그 결과 농작물은 제대로 자라지 못해 식량 부족이 심각해질 것이고, 끊임없이 발생하는 홍수 피해로 인해 인간이 안정적으로 거주할 수 있는 지역을 찾기 어려울 것이다.
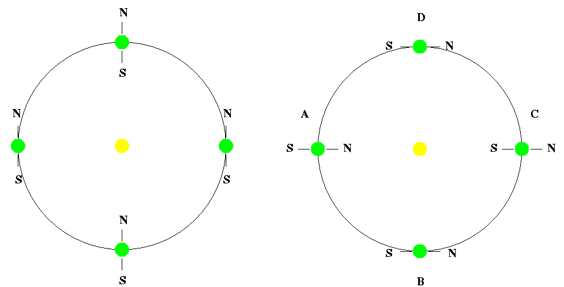
그림 2.2. 자전축이 0도인 경우와 90도인 경우
반대로, 만약 지구의 자전축이 90도로 기울어져 있다면 어떤 일이 벌어질까? 이 경우, 적도 지방에 사는 사람들은 지구가 공전 궤도상의 A 지점과 C 지점에 위치할 때 각각 한 번씩, 총 두 번의 혹독한 겨울을 겪게 되며, B 지점과 D 지점에서는 두 번의 극심한 무더운 여름을 맞이하게 될 것이다. 또한, 극 지방에 사는 사람들은 6개월 동안 낮만 계속되고, 나머지 6개월 동안은 밤만 계속되는 환경에서 살아야 한다. 북극 지방에서는 지구가 A 지점에 있을 때는 낮만 계속되는 극도로 더운 여름을, C 지점에 있을 때는 밤만 계속되는 혹독한 겨울을 맞이하게 될 것이다. 반대로, 남극 지방에서는 북극과 6개월 차이를 두고 동일한 기후 패턴이 나타나며, A 지점에서는 밤만 계속되는 추운 겨울을, C 지점에서는 낮만 계속되는 뜨거운 여름을 맞게 될 것이다.
이처럼 낮이나 밤이 지속되는 환경에서는 정상적인 경제 활동이나 사회 활동에 많은 제약이 따를 것이다. 또한, 극지방에서 극심한 추위로 인해 빙하와 바닷물이 얼어 있다가, 6개월 후 뜨거운 여름이 찾아오면 이들이 급격히 녹아 해수면이 주기적으로 상승하며, 지구상의 많은 지역이 물에 잠길 것이다. 더불어, 녹은 빙하의 차가운 물이 대량으로 바다로 유입되면서 해수 온도와 해류 패턴이 변화하고 라니냐 현상과 같은 극심한 기상이변을 발생하여 생태계의 급격한 환경 변화를 초래할 것이다.
우리 태양계의 수성과 목성의 경우, 자전축이 거의 기울어져 있지 않다(수성 0도, 목성 3도). 반면, 천왕성은 자전축이 98도로 기울어져 있다. 만약 지구의 자전축도 이처럼 수직에 가깝거나 극단적으로 기울어져 있었다면, 우리가 생명체가 살 수 있는 생명체 거주가능 영역 내에 위치하더라도, 지구는 인간이 거주하기에는 지나치게 가혹한 환경으로 변했을 것이다.
c. 적당한 자전 속도와 공전 속도를 가진 지구
인간의 생체 리듬과 이에 따른 모든 사회적 활동은 우리가 살고 있는 지구의 자전 속도에 절대적인 영향을 받는다. 지구의 자전 속도는 24시간이며, 이로 인해 낮과 밤이 각각 12시간씩 나뉜다. 이러한 주기는 하루 약 8시간 동안 일하고, 8시간은 여가 생활을 즐기며, 나머지 8시간은 수면을 취할 수 있는 이상적인 생활 패턴을 가능하게 한다. 하지만 지구와는 달리 태양계 내 다른 행성의 자전 속도는 제각각인데 우리가 이런 행성에 살고 있다면 지구와는 판이한 생체리듬을 따라야 했을 것이다.
목성에서의 자전 속도는 지구보다 짧은 10시간 정도여서 낮과 밤이 약 5시간 정도가 된다. 우리가 이런 목성의 자전 패턴에 맞게 살아가려면 약 3시간 정도는 일하고 3시간 정도는 여가 시간을 가지며 3시간 정도 잠을 자야 할 것이다. 지루하지 않아 좋을 것 같지만 현실적으로는 매우 비효율적이다. 예를 들어, 출근길 교통 체증이 심하다면 회사에 도착하기도 전에 퇴근 시간이 되어버릴 수도 있다. 집에 돌아와 잠깐 쉬려 하면 곧바로 잠잘 시간이 되고, 잠자리에 누워 조금 뒤척이다 보면 이미 아침 해가 떠 있는, 매우 불규칙하고 생산성이 낮은 행성에서의 삶이 될 것이다. 하지만 더 심각한 문제는 환경적인 변화다. 지구에서 바람이 형성되는 주요 원인은 기압 경도력, 마찰력, 그리고 지구 자전에 의한 전향력(코리올리 힘)의 상호작용 때문이다. 만약 우리가 지구보다 자전 속도가 2배 이상 빠른 행성에서 살고 있다면, 전향력이 커져 바람의 세기가 훨씬 강해질 것이다. 특히, 코리올리 힘은 극지방보다 적도 지방에서 더 강하게 작용하는데, 이러한 조건에서 태풍이 발생하면 현재보다 훨씬 더 강력한 폭풍이 형성되어 농작물 피해와 홍수 피해가 심각해질 것이다.
반대로 우리가 자전 속도가 느린 행성에서 살게 된다면 어떤 일이 일어날까? 금성에서의 자전 속도는 243일이며 낮과 밤이 약 122일 정도가 된다. 이럴 경우 태양을 향하는 면은 122일 동안 태양에 노출되어 모든것이 이글거릴 정도로 온도가 올라갈 것이면 그 반대편은 깜깜한 암흑 속에 모든 게 얼 정도로 온도가 내려갈 것이다. 이런 상황에서는 농작물 재배도 힘들어 기상재해뿐만 아니라 심각한 식량난도 초래될 것이다. 자전 속도가 느릴 경우 더욱 심각한 다른 문제가 발생한다. 지구 내부에는 금속으로 이루어진 핵이 존재하며, 지구의 자전에 의해 금속핵 내 대류 현상이 발생해 강력한 자기장이 형성된다. 그러나 자전 속도가 지나치게 느리면 대류 현상이 일어나지 않아 자기장이 발생허지 않는다. 이럴 경우 태양과 우주로부터 오는 해로운 방사선을 막아 주는 자기 방패막이 형성되지 않아 해로운 우주선에 직접 노출되어 생명체의 생존에 큰 위협을 받을 수 있다.
행성의 자전 속도뿐만 아니라 행성이 태양 주위를 한 번 도는 데 걸리는 시간인 공전 속도도 생명체의 생활 패턴에 큰 영향을 미친다. 지구의 공전 속도는 365일로 일 년에 약 3개월 단위로 봄, 여름, 가을, 겨울을 가지게 된다.
하지만 우리가 수성처럼 공전 주기가 짧은 행성에 살고 있다면 어떨까? 수성의 공전 주기는 약 88일로 봄, 여름, 가을, 겨울이 22일 주기로 바뀌게 된다. 계절이 약 3주일 주기로 바뀌게 되면 급변하는 계절에 적응하기 위해 지금보다도 훨씬 더 번거로운 삶을 살게 될 것이다. 지구상 대부분의 농작물은 파종하고 수확하는데 6개월 정도가 걸린다. 그런데 계절이 3주 단위로 급격히 변한다면 작물이 익을 시간이 충분하지 않거니와 조금 익었더라도 곧바로 온 겨울에 의해 얼어 버릴 것이다. 그럴 경우 인간의 생존에 직접적인 영향을 미치는 식량부족 사태를 야기할 것이다.
이와는 반대로 지구의 공전 주기가 해왕성처럼 164년이 되면 어떻게 될까? 이 경우 봄, 여름, 가을, 겨울의 길이가 약 40년 정도씩 되어 인간을 포함한 동식물들의 생존에 심각한 영향을 미칠 것이다. 공전 주기가 일년인 지구에서도 평년보다 겨울이 길어지면 많은 동식물이 혹독한 추위와 식량 부족으로 인해 생존에 어려움을 겪는데, 만약 겨울이 40배나 길어진다면 많은 동물이 멸종할 것이다. 인간 역시 40년 동안 농작물을 재배할 수 없다면 식량 부족으로 인해 생존 자체가 위협받게 될 것이다.
d. 적당한 크기의 지구
지구의 크기도 지구상에 살고 있는 인간의 생존과 밀접한 연관을 가지고 있다. 우리가 이 지구상에 살면서 한 번도 깊이 생각해 보지 않은 지구의 크기가 우리 인간의 생존에 중요한 영향을 미친다는 사실이 생소하게 들릴지 모르겠지만 지구의 크기는 인간뿐만 아니라 지구상의 모든 생명체에 큰 영향을 미친다. 그럼, 지구의 크기가 생명체에 어떤 영향을 미치는지 한번 알아보도록 하자.
만일 지구가 지금보다 반으로 작아진다면 어떤 일이 벌어질까? 이럴 경우 지구의 질량은 반경의 세제곱에 비례하므로 1/8로 줄어들고, 지표면의 중력은 질량에 비례하고 반경의 제곱에 반비례하므로 지금보다 반으로 줄어든다. 우리가 매일 숨 쉬고 있는 지구의 공기는 질량을 가지고 있기 때문에 지구 중력의 영향을 받게 된다. 따라서 지표면 근처에는 공기의 밀도가 높고 산으로 올라갈수록 공기의 밀도가 희박해진다. 그런데 지구가 반으로 작아지면 중력이 적어지고 이에 비례해 지구 탈출속도가 줄어들게 된다. 그럴 경우 공기 입자들의 자유 운동 속도가 지구 탈출 속도보다 커지는 상황이 생겨 많은 공기가 대기권 밖으로 달아나게 된다. 이렇게 되면 지구는 공기가 점차 희박해져서 공기로 호흡하는 모든 동식물은 생존할 수 없게 된다. 얼마나 심각한 문제인가? 실제로 지구의 반밖에 안 되는 화성에서는 적은 중력으로 말미암아 공기의 대부분이 우주로 달아나서 현재 남은 공기는 지구의 1% 정도밖에 안 되며, 지구 크기의 1/4밖에 안 되는 달은 화성보다 더 적은 중력으로 오래전에 공기가 우주 밖으로 다 날아가 버렸다.
중력이 적어지면 공기가 없어지는 것 외에 우리의 신체에도 변화가 일어난다. 우주 비행사들이 중력이 약한 우주 공간에 가면 관절이 늘어나듯이 사람들의 키가 약한 중력으로 인하여 지금보다 더 커지며 걸음걸이는 중력이 적은 달에서의 아폴로 우주인처럼 깡충깡충 뛰는 걸음걸이가 될 것이다. 성냥개비처럼 삐쩍 마른 사람이 토끼처럼 깡충깡충 뛰어다닌다면 얼마나 우스꽝스러울 것인가? 그런데 다행히도 창조주 하나님께서는 하나님의 형상을 따라 아름답게 창조된 우리가 이런 우스꽝스러운 모습이 되지 않도록 지구를 적당한 크기로 만들어 놓으셨다.
그와 반대로 만일 지구의 크기가 지금보다도 더 컸다면 어떻게 되었을까? 지구의 크기가 커져서 야기되는 문제 역시 중력과 연관되어 있다. 지구의 중력이 커지면 수소나 헬륨 등 가벼운 기체뿐만 아니라 몸에 해로운 암모니아나 메탄 같은 기체들이 우주로 탈출을 못 하고 대기에 점점 쌓이게 된다. 이렇게 되면 중력이 큰 목성처럼 지구의 대기 성분이 바뀌게 되는데 생명 활동에 필요한 산소보다는 몸에 해로운 메탄 같은 유독 가스들이 쌓이게 되어 아예 생존 자체가 불가능하게 된다.
지구의 크기가 커져서 생기는 또 다른 위협은 커진 중력으로 인해 지금보다도 훨씬 더 많은 운석을 지구로 끌어당겨 우리의 생존에 직접적인 피해를 입히게 된다는 사실이다. 운석이 지금보다 더 많이 떨어진다면 운석이 어디로 떨어지게 될지에 대한 운석 예보가 일기 예보보다 더 큰 비중을 차지하게 될 것이다. 외출 시에는 모든 사람이 두꺼운 헬멧을 착용해야 할 것이며 집이나 자동차들은 모두 두꺼운 철판으로 무장하여 떨어지는 운석에 대비해야 될 것이다. 또한 지구의 중력이 커지게 되면 무거운 중력을 이기고 걸어 다니기 위해 모든 사람이 코끼리 다리처럼 튼튼한 다리를 가지고 어슬렁어슬렁 걷게 될 것이며 키도 잘 자라지 않게 될 것이다.
지구의 크기 변화에 따른 산소량 변화도 인간의 생존에 큰 영향을 미친다. 만일 현재 대기의 21%를 차지하고 있는 산소의 양이 지금보다 적다면 어떨까? 그렇게 되면 당연히 숨쉬기가 힘들 뿐만 아니라 많은 산소가 요구되는 스포츠 활동은 결코 할 수 없는 제한적인 삶을 살게 될 것이다. 야트막한 야산에 올라가는데도 산소통을 지고 가야 될지도 모르는 상황이다.
이와 반대로 만일 산소가 지금보다 많이 존재한다면 어떻게 될까? 이 경우 신선한 산소를 많이 마실 수 있어 좋겠지만 많은 산소로 인하여 지구는 항상 불난리를 겪게 될 것이다. 일상생활에 있어서도 쉽게 불꽃이 발화하여 어느 공무원들보다 소방 공무원들이 더 많을 것이다. 산소가 많아서 생기는 또 다른 문제점은 산소 원자 3개가 모인 오존이 지나치게 많이 생성된다는 점이다. 오존을 호흡 중에 많이 들이켜면 우리 허파에 손상을 주어 위험하며 특히 호흡기 질환이 있는 사람에게는 치명적이다. 이런 이유로 오존 주의보가 발령되는데 오존이 지나치게 많다면 외출 시에 누구나가 다 오존을 정화해 주는 마스크를 쓰고 나가야 하는 불편한 세상이 될 것이다.
그림 2.3. 태양계 행성 크기 비교
e. 강한 자기장을 가지고 있는 지구
지구는 우주 공간으로부터 끊임없이 쏟아지는 우주선(Cosmic ray)에 노출되어 있다. 대부분은 가까운 태양으로부터 태양풍과 같이 방출되는 하전 입자들이거나 초신성의 폭발로 생긴 입자들이다. 이런 입자들이 걸러지지 않고 지구로 바로 들어올 경우 지구 생명체의 생존에 심각한 영향을 미친다.
그 첫 번째 영향은 하전 입자들의 높은 에너지로 인해 세포가 파괴되거나 돌연변이를 일으키게 되는 것이고 두 번째 영향은 이런 하전 입자들이 당구공이 다른 당구공을 밀어내듯이 때려내기 작용(Sputtering)에 의해 지구 상층 대기의 공기들을 우주 공간으로 날려 보내어 공기가 줄어들게 만든다는 것이다. 우리가 숨 쉴 수 있는 지구상의 공기를 감소시키니 얼마나 심각한 문제인가? 실제로 화성의 자기장은 지구 자기장의 만분의 일 정도가 되는데 이런 약한 자기장으로 인해 그나마 조금 남아 있는 이산화탄소로 이루어진 대기가 이 때려내기 작용에 의해 점점 더 희박해지고 있다. 그런데 다행히도 우리 지구는 이런 위험한 현상을 방지해 주는 효율적인 자기 방패막을 가지고 있다.
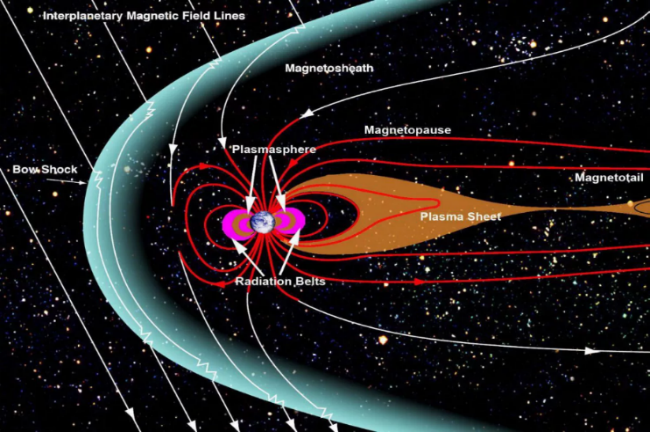
그림 2.4. 지구를 감싸고 있는 자기 방패막
지구 내부에는 철과 니켈로 이루어진 액체 상태의 금속핵이 있는데 지구의 자전으로 인해 이 금속핵이 대류 현상을 일으키게 되면 자기장이 발생하고 이렇게 생긴 지자기는 다음 그림에 나타난 것처럼 지구 전체를 감싸는 거대한 자기권을 형성한다. 태양 활동에 의해 태양풍과 함께 불어오는 하전 입자들이 지구 근처로 다가올 경우 이렇게 형성된 거대한 자기권에 의해 지구상으로 바로 들어오지 않고 자기권계면을 따라 비껴가도록 한다. 그런데 지구상에 자기 방패막이 조금 약한 곳이 있는데 그곳이 남극과 북극 지방이다. 하전 입자들이 극지방에 들어오면 상층 대기와 충돌하면서 방전을 일으키는데 그게 바로 형형색색으로 빛나는 오로라로 나타난다.
이런 자기 방패막이 없었다면 고에너지 우주선들이 여과 없이 들어와 세포를 파괴하고 지구의 공기를 우주로 탈출시켜 생명체들의 생존에 치명적인 영향을 미쳤을 것이다.
f. 예외적으로 큰 달을 가지고 있는 지구
달은 지구 생명체의 존재에 중요한 영향을 미친다. 우리가 밤하늘에서 늘 보아왔던 달이 우리의 생존과 어떤 관련이 있을지 의아하겠지만, 달은 두 가지 중요한 역할을 한다. 하나는 지구 자전축을 안정화해 기상 이변을 방지하고, 다른 하나는 조석력을 발생시켜 해양 생태계를 유지하는 것이다.
먼저 달에 의한 지구 자전축의 안정화에 대해 알아보자. 우리가 앞서 살펴보았듯이 지구의 자전축은 23.5도로 기울어져 있어 지구상의 생태계가 쾌적하게 살 수 있는 계절의 변화를 가져다준다. 그런데 지구의 자전축이 이렇게 23.5도로 일정하게 유지될 수 있는 이유는 지구 가까이에 있는 달의 존재 때문이다.
태양계 내의 지구형 행성에서 우리 지구처럼 커다란 달(반지름=1,740km)을 가진 행성은 없는데 같은 지구형 행성인 수성이나 금성에서는 아예 달이 없고 화성에는 그리스 신화에 나오는 포보스와 데이모스의 이름을 딴 2개의 달이 있지만 반지름이 달의 0.6%인 약 10km 정도로 아주 작다.
앞서 논의하였듯이 태양계에서의 생명체 거주가능 영역은 1천문단위 근처에 있다. 반면에 태양계의 행성들이 지나다니는 황도면은 해왕성까지 약 30 천문단위에 걸쳐있다. 이 거리를 고려하면 지구는 태양에 너무 가깝게 붙어있다. 이렇게 가까이 있어야만 생존에 적당한 온도를 가질수 있지만 한가지 문제가 존재한다. 그것은 지구에 미치는 태양의 중력이 너무 강해 지구의 자전축을 불안정하게 한다는 것이다.
태양의 중력에 의한 자전축 흔들림 현상은 이웃 행성들에서도 관측될 수 있다. 아래 그림은 큰 달이 없는 화성에서 태양의 중력에 의해 나타나는 자전축과 이심률의 변화를 보여준다. 그림을 보면 화성의 자전축과 이심률은 약 15만 년 주기로 크게 진동하며 과거 6백만 년 동안 자전축은 15도에서 45도까지, 이심률은 0.01에서 0.11까지 변하였음을 알 수 있다.
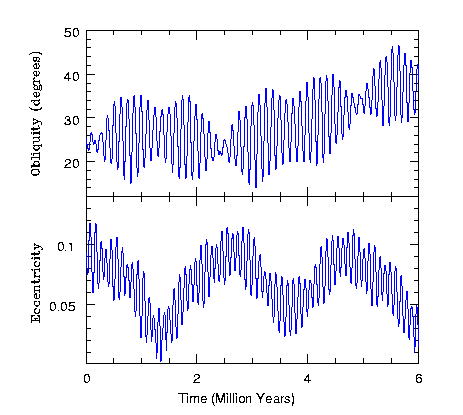
그림 2.5. 화성 자전축과 이심률의 변화
지구와 비슷한 크기의 자매 행성인 금성에서는 달이 없음으로 말미암아 태양의 중력으로 인해 자전축의 변화가 0도에서 180도로 카이오틱하게 변한다. 만일 지구에 달이 없다고 가정한면 예상되는 자전축의 변화는 어떻게 될까? 논문에 따라 다르지만 대개 수백만년 동안 약 0도에서 70도 정도의 변화를 가져온다고 한다. 이 정도로 자전축이 변화되면 심각한 기상재해를 야기하게 될 것이며 궁극적으로 생명체의 생존에 치명적 결과를 가져올수 있을 것이다. 하지만 다행히 창조주께서는 지구 곁에 예외적으로 큰 달을 준비해 놓으셔서 이런 재해를 방지 하셨다.
그렇다면 다른 지구형 행성들과는 달리, 지구는 어떻게 그렇게 거대한 달을 위성으로 가질 수 있었을까? 달의 기원에 대해서는 몇 가지 가설이 제시되어 왔다. 화성 크기의 천체가 지구와 충돌하여 달이 형성되었다는 충돌설, 다른 행성을 중력으로 붙잡았다는 포획설, 지구와 달이 동시에 형성되었다는 동시 형성설, 그리고 지구의 일부가 떨어져 나가 달이 되었다는 분열설 등이 그것이다. 그러나 최근의 연구들은 충돌설을 가장 유력한 설명으로 지지하고 있다. 그런데 거대 충돌을 통해 지구–달 시스템과 같은 대형 위성이 형성되기 위해서는 여러 조건이 정밀하게 충족되어야 한다.
첫째, 충돌체의 크기와 충돌 각도가 매우 정교하게 맞아떨어져야 한다. 충돌체(테이아)는 지구를 완전히 파괴하지 않으면서도 맨틀 물질을 우주 공간으로 충분히 분출시키기 위해, 지구 질량의 약 10% 정도 크기여야 하며 반드시 비스듬한 각도로 충돌해야 한다. 정면 충돌은 달을 형성할 재료를 만들어내지 못한다.
둘째, 분출된 잔해 물질은 빠르게 응축되고 합체되어야 한다. 잔해가 지구의 강력한 조석력에 의해 분산되거나 다시 지구로 흡수되기 전에, 짧은 시간적 창(accretion window) 안에 달을 완성해야 한다.
결정적으로, 달의 안정적인 형성을 위해서는 충돌 직후 지구의 자전 속도가 충분히 빨랐어야 한다. 왜냐하면 충돌로 분출된 물질이 지구로 다시 떨어지지 않고 안정된 궤도를 형성하여, 로슈 한계 바깥에서 달로 응집되어야 하기 때문이다. 이처럼 크기, 각도, 속도, 시간적 조건이 모두 완벽하게 맞아떨어져야 하므로, 다른 외계 지구형 행성에서 이처럼 거대 위성이 형성되는 것은 극히 드문 현상일 것으로 추정된다.
이제 달의 조석 현상이 해양 생태계에 어떤 영향을 미치는지 알아보자. 달의 조석 현상은 밀물과 썰물을 발생시키며 이에 따른 해류의 움직임을 발생해 갯벌에서 생활하는 어패류에 풍부한 산소와 영양분을 전달한다. 또한, 해류의 움직임은 해양 생태계 먹이사슬 구조의 가장 기본인 플랑크톤에 풍부한 산소를 공급해 주고 영양분이 풍부한 하층의 영양염을 상층으로 퍼 올리는 용승류를 발생시켜서 플랑크톤에게 영양을 제공하며 유영력이 없거나 미약한 플랑크톤을 여러 곳으로 퍼트려 많은 물고기의 먹이로 공급해 주는 역할을 한다.
밀물과 썰물의 또 다른 중요한 역할은 강에서 흘러온 영양분이 많은 민물을 바닷물과 섞는 역할을 한다. 달에 의한 조석 간만의 차이가 없으면 염분 농도가 높은 무거운 바닷물은 아래로 가라앉고 강에서 유입되는 영양분이 풍부한 가벼운 물은 상층에 떠 있는 성층화 현상이 일어난다. 이런 성층화가 일어나면 상층에 집중된 영양분을 먹이로 하는 적조 플랑크톤이 대량 번식하여 다음 그림에 나타난 것처럼 적조가 발생한다. 또한 그 맹독성 적조를 먹은 어패류는 폐사되며 대량으로 번식된 플랑크톤의 잔재들이 바다 밑바닥에 쌓이면 세균들이 이들을 분해하면서 대부분의 산소를 고갈시키게 된다. 대량 번식된 플랑크톤들은 물고기의 아가미에 붙어서 물고기를 질식시키기도 하며 편모조류 플랑크톤인 코콜리디니움은 독을 내뿜어 물고기를 죽이기도 한다.

그림 2.6. 적조현상에 의해 붉게 변한 바다
또한 조석 현상이 없으면 그나마 조금 남아 있는 산소를 포함하고 있는 상층부의 물이 아래층으로 전달되지 않아 그곳에 살고 있는 어류들의 대량 사멸을 불러오는데 이럴 경우 육지 근처에서 살고 있는 작은 물고기를 먹이로 하는 큰 물고기들도 먹이가 없어져 궁극적으로 해양 생태계의 대붕괴를 가져올 수 있다. 여름철 장마로 많은 비가 내려 강물이 바다로 유입되면 그 후 적조 현상이 일어나는 것도 이 성층화 현상 때문인데 조석 간만의 차이가 큰 곳이나 물살이 빠른 해안에서는 강물과 바닷물이 잘 섞여 그나마 적조 현상이 적게 생기지만 섬들이 많아 물의 흐름이 대체로 느린 해안에서 적조 현상이 많이 일어나는 것도 이와 같은 이유 때문이다.
이렇듯 달은 지구의 자전축을 안정시키고 해양 생태계를 유지하는 중요한 역할을 한다. 그러나 만약 지구가 달을 가지고 있더라도 현재의 달보다 크기가 작았다면 지구 자전축의 안정화 효과가 충분하지 않았고 조석력도 적었을 것이다. 반대로, 달이 지금보다 더 컸다면 지구와 달 사이의 중력이 강해져 달이 지구의 중력에 끌려와 충돌했을 가능성도 있다. 또는, 커진 달의 강한 중력으로 인해 조석력이 증가하여 해안가의 많은 도시들이 밀물 때마다 물에 잠기는 상황이 발생했을 것이다. 그뿐만 아니라 현재와 같은 크기의 달이 있더라도 지금보다 더 가까운 거리에 위치하거나 또는 더 멀리 떨어져 있었다면 유사한 문제들이 발생하였을 것이다. 이렇게 우리의 생존에 중요한 달도 하나가 아니라 두 개였으면 또 다른 위협으로 다가왔을 것이다. 증가한 조석력으로 인해 수십에서 수백 미터 높이의 파도가 형성되고, 그로 인한 거대한 쓰나미가 발생했을 가능성이 크다. 또한, 강한 중력의 영향으로 지구의 화산 활동과 지진이 증가하여 험난한 자연환경을 가졌을 것이다.
g. 목성에 의해 보호받고 있는 지구
목성은 태양계의 가장 큰 행성이며 지구보다도 11.2배 크고 무게는 318배 무겁다. 목성의 존재가 우리의 생존에 어떤 연관이 있을까 하고 궁금해하겠지만 목성 역시 달과 못지않게 우리의 생존에 중요한 역할을 한다. 우리 태양계에는 화성과 목성 사이 그리고 목성 궤도 근처에 소행성대가 존재하는데 궤도를 이탈한 소행성들과 주기적 또는 비주기적으로 태양을 방문하는 혜성들이 지구에 큰 위협을 줄 수 있다. 이런 소행성들이나 혜성 또는 이들의 부서진 파편들이 지구상에 떨어지는 것을 운석이라고 부르는데 지금 이 시간에도 수많은 운석이 지구 대기권으로 떨어지고 있다. 아래 그림은 지구로 떨어지는 운석의 크기와 그러한 운석이 얼마나 자주 떨어지는가를 보여 주는 그림이다.
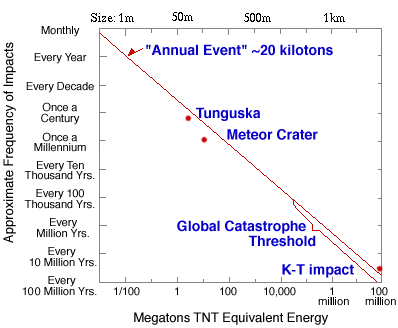
그림 2.7. 지구상에 떨어지는 운석의 크기와 빈도수
직경 1미터 정도인 운석은 한 시간에 하나 정도로, 수 미터 크기의 운석은 매일 한 번 정도 그리고 집채만 한 수 미터에서 10미터 크기의 운석은 일 년에 한 번 정도로 지구상에 떨어진다. 대기권으로 진입하는 운석의 직경이 10미터 이하일 경우 대부분은 공기와의 마찰로 인해 대기 중에서 다 타 버려 실제로 지표상에 어떤 충격을 주지 못하지만 문제는 그보다 더 큰 운석이 지구로 떨어질 경우이다.
이러한 운석들이 지표면에 떨어지면 심각한 재난을 초래한다. 실제로 1908년 소련 퉁구스카 지역에서 약 60미터 크기의 운석이 떨어지면서 공중에서 폭발하였는데 그 충격에 의해 약 2천km2 면적의 지역에서 약 8천만 그루의 나무가 쓰러졌다고 한다(그림 참조). 이런 크기의 운석은 1세기에 한 번 정도로 떨어지며 그 폭발력은 히로시마 원폭 1,000개에 해당한다. 100미터 크기의 운석은 천 년에 한 번 정도, 1km 크기의 운석은 백만 년에 한 번 정도에 그리고 백악기 말인 6,500만 년 전에 멕시코의 유카탄반도에 떨어져 지구상의 공룡들을 멸종시킨 약 10km 크기의 운석은 1억 년에 한 개 정도가 떨어진다. 100m 크기의 운석이 떨어지면 약 1km 정도의 운석 구덩이가 생기며 수백 메가톤의 TNT를 터트린 것과 같은 충격을 미친다. 그럴 경우 반경 15km 내의 땅을 초토화시키고 만일 그 운석이 바다에 떨어질 경우 수십 미터 높이의 쓰나미로 인해 해안가에 있는 대부분의 도시가 물에 휩쓸리게 된다.

그림 2.8. 퉁구스카 운석에 의해 쓰러진 나무들
목성이 지구상의 생명체에 중요한 이유는 이렇게 지구에 큰 위협을 주는 운석들을 지구 대신에 끌어들여 지구를 지켜주는 방패 역할을 한다는 데 있다. 계산에 의하면 직경이 1km 이상의 주기 혜성들을 목성이 지구보다 약 5,000배 정도나 잘 포획하는데 만일 지구 가까이에 목성이 없었다면 이런 혜성들이 목성 대신에 지구에 떨어져 엄청난 피해를 초래하였을 것이다.
목성의 강한 중력이 실제로 혜성을 포획하는 장관이 1994년에 벌어졌다. 다음 그림은 목성의 중력에 의해 여러 조각으로 부서진 슈메이커-레비 혜성과 이 혜성의 부서진 조각들을 목성이 진공청소기처럼 빨아들여 목성 표면에 부딪치는 장면을 허블 우주 망원경이 관측한 사진이다. 사진에 갈색으로 보이는 부분들이 혜성 조각이 충돌한 지점인데 충돌한 혜성 중 가장 큰 조각은 지름이 2km 정도로 만일 이것이 목성 대신 우리 지구에 떨어졌다면 북미 대륙 정도의 땅을 초토화했을 정도의 위력이었다.

그림 2.9. 부서진 슈메이커-레비 혜성과 목성에 충돌한 모습
그런데 한 가지 중요한 사실은 목성이 지금보다 지구로부터 더 멀리 떨어져 있었거나 아니면 지금의 크기보다 조금 더 작았다면 지구에 이렇게 큰 위협을 줄 수 있는 많은 운석을 잘 포획하지 못했을 것이다. 적당한 크기의 목성이 지구로부터 적당한 거리에 존재하여 지구를 위험한 천체들로부터 지켜 준다는 것이 경이롭고 다행스러운 일이다.
h. 판구조론에 의해 자동 온도 조절이 되는 지구
판구조론(Plate tectonics)은 지각을 구성하고 있는 판들이 지구 맨틀의 대류현상으로 인해 움직이며 이에 따라 지진과 화산활동이 일어나며 습곡과 산맥 등이 형성된다는 설이다. 그렇다면 이 판구조론이 우리의 생존과 무슨 연관이 있을까? 결론부터 말하면 판구조론은 지구의 온도를 자동으로 조절하여 지구상의 물이 얼거나 또는 끓지 않도록 해 준다.
그림 2.10. 지각을 구성하고 있는 여러 판들
판구조론이 어떻게 지구의 자동 온도 조절기 역할을 하는지 알아보자. 지구의 온도는 여러 요인에 의해 결정되는데 그중 가장 중요한 것들은 태양으로부터 지구에 도달하는 태양 에너지의 세기와 그 태양 에너지가 실제로 지구상에 흡수되는 양 그리고 지구 대기의 수증기, 구름, 이산화탄소에 의한 온실 효과에 달려 있다. 태양에서 도달하는 빛은 지구상의 대기를 잘 통과하지만 일단 지표면에 한 번 반사되면 난반사가 되고 파장이 길어져 대기 중의 온실효과 가스에 흡수된 후 반은 우주로 방출되고 반은 지구상으로 방출되어 지구가 더워진다.
온실효과를 일으키는 가스들은 이산화탄소, 메탄, 오존 등이 있는데 가장 큰 영향을 미치는 것은 이산화탄소이다. 대기 중에 이산화탄소가 많을 경우 온실효과가 커져서 지구 전체의 온도가 올라가고 적을 경우에는 온실효과가 적어 지구의 온도가 내려간다. 이산화탄소의 많은 부분은 화산 활동에 의해 분출된다. 이렇게 분출된 이산화탄소는 풍화작용에 의해 생성된 규산염 광물과 반응하여 탄산칼슘(주로 석회석)을 생성하면서 대기 중에서 제거된다. 그런데 대기 중에 이산화탄소가 많아 지구의 온도가 올라가면 규산염 광물의 풍화작용도 함께 상승한다. 이렇게 풍화작용이 상승하면 더 많은 규산염 광물이 생성되어 대기 중의 이산화탄소와 많이 반응하여 석회석을 만들면서 이산화탄소량을 감소시키고 그렇게 되면 감소한 이산화탄소로 인한 온실효과도 감소하어 지구의 온도가 내려간다. 이렇게 하여 지구의 온도가 내려가면 규산염 광물의 생성도 함께 감소하며 이산화탄소와의 반응이 작아져 대기 중에 이산화탄소의 비율이 올라가고 온실효과도 올라가면서 지구의 온도가 점점 상승한다. 지구의 온도가 상승하면 또다시 이와 같은 사이클을 거쳐 지구의 온도를 언제나 적정 상태로 유지하게 된다. 즉, 이산화탄소의 순환에 의해 지구의 온도가 자동 조절된다는 이야기다. 아래 그림은 지난 80만 년 동안 이 과정이 어떻게 작동되었는지를 보여 주는 그림이다. 이산화탄소량이 증가하면 지구의 온도가 증가하고 이산화탄소가 감소하면 지구의 온도가 감소함을 보여 주고 있다.
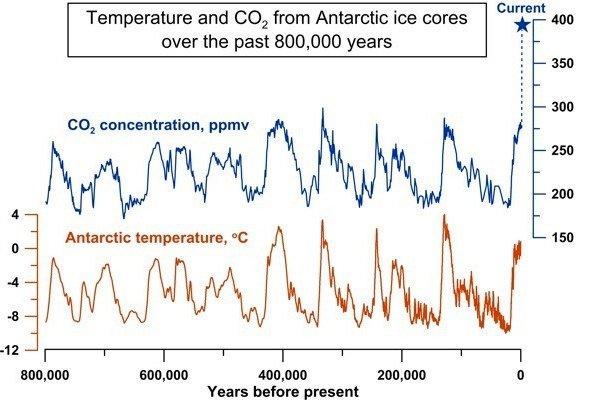
그림 2.11. 이산화탄소 증가량과 지구 온도의 상관 관계
그런데 놀라운 것은 이산화탄소에 의한 자동 온도 조절 작용이 판구조론에 의한 지각의 이동이 없다면 작동하지 않는다는 사실이다. 다음 그림처럼 지구는 화산활동을 통해 이산화탄소를 대기 중으로 분출하고 이렇게 분출된 이산화탄소는 풍화작용으로 생성된 규산염 광물과 반응하여 석회석으로 변해 바닷속의 침전물 형태로 퇴적된다. 바닷속에 퇴적된 석회석은 해저 지각의 한판이 다른 판 아래로 들어가는 섭입(subduction) 작용에 의해 지구 내부로 들어가 뜨거운 열에 의해 녹으면서 마그마 분출과 함께 이산화탄소를 다시 대기 중으로 방출한다.
만일 판구조론에 의한 섭입 작용이 없다면 바닷속에 퇴적된 석회석은 계속 쌓여 바닷속에만 머물게 되고 그렇게 되면 이산화탄소가 대기 중으로 방출되지 않아 온실효과가 줄어들면서 지구의 온도가 급격히 내려가게 된다. 만일 역사상 한 번이라도 이산화탄소 순환 사이클이 깨어져 작동하지 않았다면 지구상에 생명체가 존재하기 힘들었을 것이다. 왜냐하면 온도가 내려가 바닷물이 얼 정도까지 되면 태양으로부터 오는 대부분의 빛은 지구에 흡수되지 않고 얼음에 반사되는데 이렇게 되면 더 이상 돌이킬 수 없는 ‘폭주 빙실 효과(runaway icehouse effect)’에 의해 지구가 얼음에 덮인 행성으로 변할 것이기 때문이다.
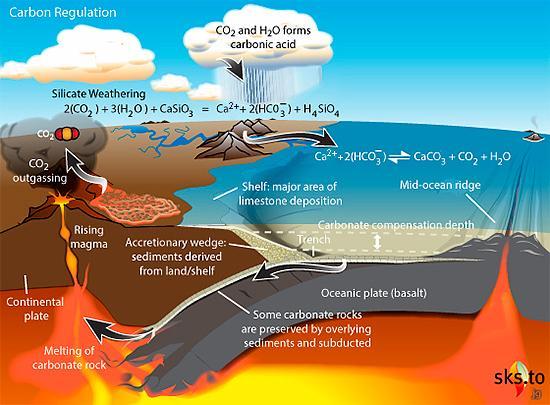
그림 2.12. 섭입 작용에 의한 이산화탄소 순환 과정
지구 자동 온도 조절에 중요한 역할을 하는 이 판구조론이 어떻게 시작되었고 또한 오랜 기간 동안 어떻게 유지되었는가에 대한 연구가 극히 최근에야 이루어지고 있다. 연구가 거듭될수록 판구조론에 대한 신비로운 사실이 밝혀지고 있는데 최신 연구 결과에 의하면 만일 지구의 크기가 현재보다 20% 정도 크거나 또는 적거나, 지각에 포함된 철이나 니켈 등의 금속 성분이 지금보다 조금 많거나, 또는 지각의 두께가 지금보다도 더 두꺼웠다면 판구조론이 정상적으로 작동되지 않았다고 한다.
지구가 적당한 두께의 지각을 가지고 있고 그 지각이 판구조론에 의해 서서히 이동하여 대기 중에 0.03%밖에 되지 않는 이산화탄소를 순환시켜서 지구의 온도를 자동으로 조절되게 하는 것이 얼마나 신비스러운가?
i. 적당한 크기의 태양을 가지고 있는 지구
우리 태양계에 있어 물이 얼지도 않고 끓지도 않은 영역을 생명체 거주가능 영역이라고 하였는데 이 생명체 거주가능 영역은 우리가 어떤 별 주위에 사느냐에 따라 그 영역이 달라진다. 다음 그림에 나타난 것처럼 태양보다 작은 별 주위에 살면 별의 복사열이 적어 생명체 거주가능 영역은 그 별에 더 가까이 위치하게 되고 태양보다 큰 별 주위를 산다면 많은 복사열 때문에 생명체 거주가능 영역이 그 별에서 더 멀어지게 된다.
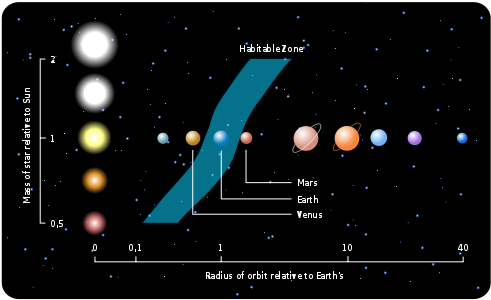
그림 2.13. 별의 크기에 따른 생명체 거주가능 영역의 변화
별의 질량이 적어 생명체 거주가능 영역이 그 별에 가까이 위치할 경우 지구와 달의 경우처럼 조석 잠김 현상이 생기는데 그렇게 되면 그 별과 행성이 동주기 자전(Synchronous rotation)을 하게 된다. 양손을 맞잡고 마주 보면서 도는 것처럼 동주기 자전하게 되면 별을 마주 보고 있는 쪽만 항상 햇빛을 받게 되므로 물이 끓을 정도의 온도가 되고 그 반대쪽은 항상 밤이고 모든 게 얼어붙은 동토의 땅이 된다. 따라서 이런 환경의 행성에서는 생명체의 생존은 거의 불가능하다. 또한 행성이 자전하지 않기 때문에 자기장을 형성할 수가 없다. 자기장이 없다면 별에서 나오는 유해한 방사선이 세포와 DNA를 손상해 생존을 위협한다.
반대로 별의 질량이 커서 생명체 거주가능 영역이 별에서 멀리 떨어져 있으면 생명체 거주가능 영역의 수명이 오래가지 못한다는 단점이 있다. 왜냐하면 질량이 큰 별들은 질량이 작은 별들에 비해 핵연료인 수소를 더 빨리 소모하게 되고 헬륨 연소를 시작하면서 적색거성으로 진화하는데 이렇게 되면 별의 크기가 점점 커져서 근처에 있는 행성 자체를 삼킬 정도가 된다. 적색거성으로 진화하는 별의 수명은 대개 수백만 년에 불과하며 DNA와 세포에 해로울 수 있는 높은 수준의 자외선과 X-선을 방출하여 생명체가 살기 어려운 환경을 만든다. 또한 별의 에너지 방출에 많은 변동이 있어 행성의 기후를 불안정하게 만들고 극심한 온도 변화를 일으켜 생명체의 생존을 어렵게 만들 수도 있다.
따라서 오랫동안 안전한 생명체 거주가능 영역에 머물기 위해서는 태양과 비슷한 크기의 별을 지구의 에너지원으로 가지고 있어야 한다. 그럴 경우 적색 왜성이나 적색 거성과는 달리 오랜 기간 동안 안정적인 에너지를 제공하게 된다. 별들이 어떤 질량 분포를 보이는지 관측한 데이터를 보면 별들의 약 88%가 태양보다 가벼운 별들이며 태양 질량의 1-2배 사이에 있는 별들은 8%이고 나머지 4%는 태양보다 2배 이상 무거운 별들이다. 따라서 태양과 같은 질량을 가진 별들을 발견할 확률은 단지 몇 퍼센트 이내로 그렇게 높지 않음을 알 수 있다.
우리가 오랫동안 안정적인 생명체 거주가능 영역에 머물기 위해서 태양과 비슷한 질량의 별을 우리의 에너지원으로 가져야 하는 사실 외에 또 하나의 조건이 추가되는데 지구가 쌍성계가 아닌 단성계에 존재해야 한다는 것이다. 우리 인간에게도 쌍둥이가 가끔 태어나지만 태양 질량의 별들에서는 거의 40% 정도가 쌍성계이다. 쌍성계는 근접 쌍성계(Close binary system)와 원격 쌍성계(Wide binary system)로 구분할 수 있다. 근접 쌍성계에서 행성이 돌고 있을 경우는 다음 그림의 상단과 같이 두 별 주위를 돌고 원격 쌍성계의 경우에는 그림의 하단처럼 한쪽별 주위나 아니면 각각의 별 주위를 돌고 있을 것이다. 만일 우리가 살고 있는 지구에 태양이 둘이라고 가정해 본다면 우리 생명체에 해로운 영향이 많을 것이다. 두 개의 태양으로 인해 우리 지구에 해로운 방사선들이 더 많이 쏟아질 것이고 두 태양의 상대적 위치에 따라 지구의 자전축과 이심률이 항시 변할 것이다. 그렇게 되면 심각한 기후 및 환경 변화를 초래되어 생명체가 살기에 적당한 행성이 되지 못하였을 것이다.
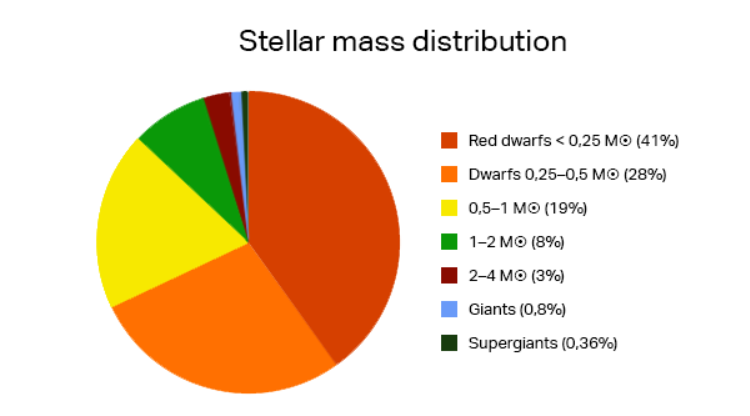
그림 2.14. 별의 질량 분포
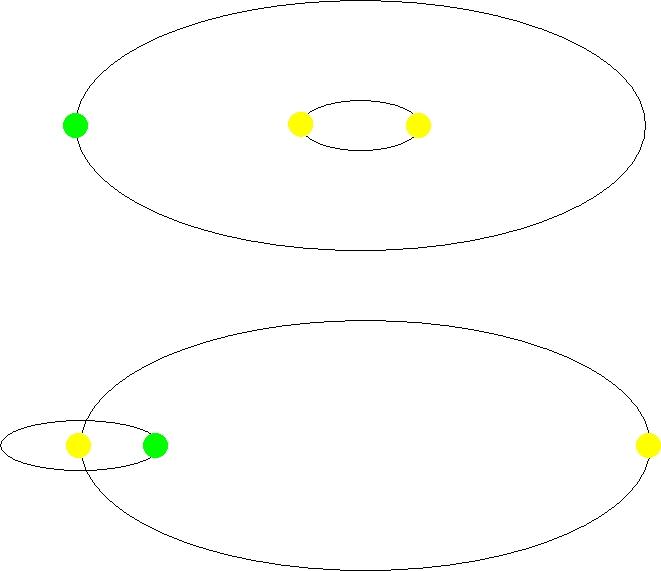
그림 2.15. 근접 쌍성계(상)와 원격 쌍성계(하)
j. 은하 중심으로부터의 적당한 거리에 위치한 지구
앞에서 보았듯이 별 주위의 특별한 영역에서만 생명체 거주가능 영역이 존재하듯이 우리 은하계 내에서도 특별한 영역에만 생명체 거주가능 영역이 존재하는데 이 영역을 은하 생명체 거주가능 영역(Galactic habitable zone)라고 부른다.
태양계 내에서의 생명체 거주가능 영역은 생명에 필수적인 물이 액체 상태로 존재하는 영역으로 정의되지만, 은하계 내에서의 생명체 거주가능 영역은 생명체와 지구를 구성하고 있는 중원소 함량이 적당하게 있는가 그리고 초신성이나 감마선 폭발의 위험으로부터 얼마나 안전한 영역에 자리 잡고 있는가 하는 문제와 연관되어 있다. 천문학에서 정의되는 중원소란 수소와 헬륨보다 무거운 원소들을 지칭하는데 이런 중원소들의 함량이 은하 내에서 균일한 분포를 보이는 것이 아니라 은하 중심에서부터의 거리에 따라 다른 분포를 보인다.
어떤 생명체가 유지되기 위해서는 섭취되는 음식물에 그 생명체의 구성 성분에 필요한 영양분이 함유되어 있어야 하고 또한 섭취되는 음식물은 흙으로부터 그 음식물 그 자체가 자라는 데 필요한 영양분을 취할 수 있어야 한다. 따라서 우리 지구를 구성하는 물질은 지구상의 생명체가 필요로 하는 유기물을 반드시 함유하고 있어야 하는데 이 유기물질을 구성하고 있는 구성 성분 대부분은 탄소, 질소, 산소와 같은 중원소들이다. 그런데 우리 태양계의 위치가 현재의 위치보다 은하 중심으로부터 좀 더 멀리 떨어져 있었다면 중원소 함량이 부족하여 생명체가 유지되지 못했을 것이고 반대로 은하 중심으로 더 가까이 있었다면 너무 많은 중원소 함량에 의해 생명체의 존재에 역효과를 가져왔을 뿐 아니라 지구의 판구조론이 잘 작동되지 않았을 것이다. 이런 이유로 해서 은하계 내에서의 태양계 위치는 우리 생명체의 존재에 있어 중요한 연관성을 가지고 있다.
우리 지구에 적당량의 중원소가 있어야 하는 또 다른 이유는 지구 내부에 존재하는 방사성 동위 원소의 함량과 연관이 있다. 지구는 태양으로부터 오는 열에너지뿐만 아니라 지구 내부에 토륨과 우라늄 같은 방사성 동위 원소들의 붕괴로 발생하는 열 에너지에 의해 데워지고 온도를 유지한다. 만일 지구 내부에 이런 방사성 동위 원소들이 없었더라면 지구는 이미 오래전에 식어서 생명체가 존재하는 행성이 되지 못하였을 것이다. 이와는 반대로 지구 내부에 너무 많은 방사성 동위원소가 존재하였다면 강한 방사선은 물론 지나치게 많은 열에너지 발생에 의해 바닷물이 끓어 생명체가 살 수 없었을 것이다. 이런 중요한 역할을 하는 방사성 동위원소의 양도 은하 내의 중원소 함량과 같은 분포를 보이는데 만일 지구의 위치가 은하 중심에서부터 외각 쪽으로 더 멀리 떨어져 있었다면 지각 내의 방사성 동위 원소량이 너무 적어 지구가 식었거나 반대로 은하 중심 쪽으로 너무 가까이 있었다면 방사선 동위 원소량이 지나치게 많아 지구의 온도가 너무 높아져서 생명체가 살 수 없는 행성이 되었을 것이다.
우리 지구가 지금보다 은하 쪽으로 좀 더 가깝게 위치했을 때 생기는 또 다른 문제점은 은하 중심으로 갈수록 증가하는 많은 별로 인해 발생한다. 지구가 속한 태양 주위에 별들이 많을 경우 그 별들에 의한 중력 섭동으로 지구의 쾌도나 자전축이 흔들리거나 심하게는 지구가 태양계 밖으로 튕겨 나갈 위험도 있다. 인근 별들의 중력 섭동은 오르트 구름과 카이퍼 벨트에 있는 천체들의 궤도를 교란시켜 더 많은 수의 혜성이 태양계로 유입되게 하며 지구와 충돌할 가능성을 높이게 된다. 또한 은하 중심은 별의 밀도가 높아 근거리에서 발생하는 초신성 폭발이나 감마선 폭발로 인한 감마선이나 엑스선 역시 지구상의 생명체들에 심각한 위협을 주게 된다. 만일 우리 태양계로부터 약 1만 광년 이내에서 감마선 폭발이 일어난다면 상대론적 속도의 고에너지 입자들이 지구상으로 쏟아져 들어와 우리 지구를 감싸고 있는 오존층을 이온화시켜 산화질소 화합물로 전환하면서 약 40%의 오존층을 파괴한다. 오존층이 파괴될 경우 태양으로부터 오는 자외선(UVB)이 걸러지지 않고 대기를 통해 들어와 우리 인간을 포함한 모든 생명체의 DNA를 파괴하여 돌연변이나 피부암을 유발한다. 40% 정도의 오존층이 파괴되면 DNA의 손상률이 현재보다 약 16배까지 증가하며 해양 생태계 먹이 사슬의 기본을 형성하고 있는 식물성 플랑크톤(Phytoplankton)을 다 멸종시킬 수 있는 양 이상의 자외선이 쏟아진다.
그림 2.16. 식물성 플랑크톤
이 식물성 플랑크톤이 없어질 경우 해양 생태계는 붕괴할 수 있으며 육상 생물들도 시간이 지남에 따라 해로운 자외선의 영향으로 점점 더 개체수가 감소되어 갈 것이다. 최근 연구 결과에 의하면 이 식물성 플랑크톤이 지구 온난화의 주범인 이산화탄소 순환에 있어서 아주 중요한 역할을 한다는 게 밝혀졌다. 식물성 플랑크톤의 광합성 작용에 의해 제거되는 이산화탄소의 양은 육지에 있는 모든 나무와 풀들이 제거하는 이산화탄소량만큼 제거한다는 것이다. 이렇게 제거된 이산화탄소는 바다 깊은 해저에 저장되는데 만일 강한 자외선으로 식물성 플랑크톤이 멸종된다면 대기 중의 탄소량이 급격히 늘어 심각한 온실효과를 일으켜 지구의 온도가 급격히 올라갈 것이다. 이렇게 심각한 영향을 미치는 감마선 폭발이 실제로 지구 역사상에 일어난 사건이 있었는데 지금으로부터 약 4억 5천만 년 전 오르도비스기 말에 해양 생태계 약 60%를 멸종케 한 사건이 지구로부터 6,000광년 떨어진 곳에서 일어난 감마선 폭발에 기인한다는 연구 결과가 발표되었다.
앞에서 언급한 것처럼 중원소 함량이 너무 많지도 적지도 않고 초신성이나 감마선 폭발의 위협을 적게 받는 은하 생명체 거주가능 영역의 범위를 계산한 논문이 사이언스지에 발표되었는데 그 영역은 은하 중심으로부터 약 2만 3천 광년부터 2만 9천 광년 사이로 추정되었다. 그런데 공교롭게도 우리 지구가 있는 태양계는 은하 중심으로부터 2만 6천 광년 떨어져 있는데 이 거리는 은하 생명체 거주가능 영역의 정중앙이 되는 위치이다. 그런데 우리가 태양계의 회전 속도와 은하의 회전 속도가 일치하는 동자전 반경(Co-rotation radius)을 고려하면 은하 생명체 거주가능 영역은 다음 그림의 녹색 영역이라고 할 수 있다. 왜냐하면 이 반경 내에 있는 별들만이 은하 회전에 의해 발생하는 밀도파에 휩쓸리지 않고 안정된 일생을 구가하기 때문이다.

그림 2.17. 은하 생명체 거주가능 영역
이 장에서 우리는 지구만이 가진 10가지 특별한 조건들을 통하여 인간을 비롯한 다양한 동식물들이 생존할 수 있는 절묘한 환경들을 살펴보았다. 이러한 10가지의 환경 중 어느 한 가지라도 어긋났더라면 지구는 인간이 정상적인 삶을 영위할 수 없는 극한 환경이 되었거나 생명체가 살 수 없는 행성으로 남았을 것이다. 그리고 이러한 조건들은 아주 정교하게 조정이 되어있어 우연히 생성되었을 확률은 극히 희박함을 알 수가 있다. 이는 지구가 단순한 우연의 산물이 아니라 생명체의 생존과 보존을 위하여 창조주에 의해 설계된 아주 특별한 행성임을 시사한다. 다음 장에서는 우리의 존재 자체가 단순한 우연의 결과인지 아니면 하나님의 창조 섭리에 의한 것인지 좀 더 자세히 알아보도록 하자.
3. 창조인가 진화인가?
생명체는 지구상에 어떻게 나타났는가? 단순한 우연에 의한 진화의 결과인가, 아니면 창조의 결과인가? 이 장에서는 진화론과 창조론에 관한 다양한 주제를 다루면서 창조가 맞는지 아니면 진화가 맞는지 살펴보려고 한다. 먼저, 생명체가 지구상에 어떻게 나타났는지에 대한 생명의 기원을 자세히 살펴보고, 다윈의 이론이 '진화론'으로 불려야 하는지 아니면 '유전적 적응론'이라고 불러야 하는지 알아볼 것이다. 그 후, 인간이 유인원으로부터 진화했는지, 지적설계론은 무엇인지 알아보고, 입자 물리학, 외계 생명체의 존재 가능성, 동물의 본능, 그리고 자연에 나타나는 수학적 원리 등을 통해 진화론의 타당성을 살펴볼 것이다.
a. 최초의 생명체는 지구상에서 어떻게 나타났는가?
최초의 생명체가 지구상에 어떻게 나타났는가에 따라 창조론과 진화론의 진위가 결정된다. 생명체가 자연적으로 나타났다면 진화론이 맞을 것이고 창조주의 개입 없이는 결코 생겨날 수 없다면 창조론이 맞을 것이다.
모든 생명체는 세포로 이루어져 있으며, 세포는 핵산(DNA·RNA), 단백질, 지질, 탄수화물 등으로 구성되어 있다. 단백질은 아미노산이 펩타이드 결합으로 중합되어 만들어지며, 아미노산은 DNA에 기록된 유전 정보를 바탕으로 만들어진다. 지질과 탄수화물 또한 DNA의 유전정보로 만들어진 효소에 의해 만들어지며, 이 효소들 역시 단백질로 구성되어 있다. 따라서 생명체의 기본 단위인 세포가 형성되기 위해서는 단백질이 필수적이며, 단백질이 형성되기 위해서는 DNA와 RNA가 이미 존재해 있어야 한다. 그런데 문제는 초기 지구에 DNA, RNA, 단백질, 효소 등이 전혀 존재하지 않았다는 점이다. 즉, 생명체의 시작 자체가 자연적으로 발생하기가 전혀 불가능하다는 것이다.
그럼에도 불구하고 진화론자들은 초기 지구에서 RNA가 먼저 스스로 형성되어 전령 RNA(mRNA)로서 유전 물질의 역할을 70하고, 리보솜 RNA(rRNA)로서 단백질 합성에 관여하는 리보자임 효소 역할도 하며, 단백질과 DNA를 만들어 냈다는 ‘RNA 월드 가설’을 주장한다.
하지만 실제로는 RNA 분자 자체가 극히 복잡하고 정교한 서열을 요구하여 무작위적으로 자연 생성될 확률이 사실상 zero에 가깝고, 설령 우연히 만들어졌다 하더라도 화학적으로 매우 불안정하여 금세 분해되며, 본질적으로 RNA가 수행할 수 있는 촉매 기능은 진정한 단백질 효소에 비해 극히 제한적이고 효율이 낮다. 게다가 단백질이 없으면 RNA가 만들어질 수 없으며, RNA가 없으면 단백질이 만들어질 수 없기 때문에 RNA 월드 가설은 구조적 모순을 안고 있다.
더 나아가 설령 RNA, 단백질, DNA가 우연히 모두 만들어졌다 하더라도, 이것들이 스스로 조립되어 살아있는 원핵세포를 이루고, 이어서 진핵세포로 진화하며, 세포소기관이 국재화되고, 세포 분화가 일어나며, 조직·기관이 형성되고, 최종적으로 다세포 생물이 출현하는 모든 단계는 각각 천문학적 확률의 연속이다.
이제 생명체의 자연발생 가능성을 본격적으로 논하기 위해, 먼저 생명 기원 연구에서 자주 언급되는 밀러-우레이 실험을 살펴본 뒤, RNA의 형성, 단백질의 형성, DNA의 형성, 원핵세포의 형성, 진핵세포의 형성, 세포소기관의 국재화, 세포 분화, 조직과 기관의 형성, 그리고 다세포 생명체의 형성 등 일련의 주제들을 차례로 탐구해 보겠다.
i. 밀러-우레이 실험
밀러-우레이 실험은 원시 지구 환경에서 아미노산 등 생명 구성 유기물이 무기물로부터 자연적으로 합성될 수 있는지를 확인하기 위해 수행된 실험이다.
이 실험에서 그들은 지구의 원시 대기가 메탄, 암모니아, 수소, 수증기로 이루어졌다고 가정한 후, 이 혼합 기체를 가열 장치, 진공 펌프, 방전 장치, 냉각 장치 등으로 구성된 실험 장치에 넣어 반응시킨 결과, 네 종류의 아미노산을 얻었다.
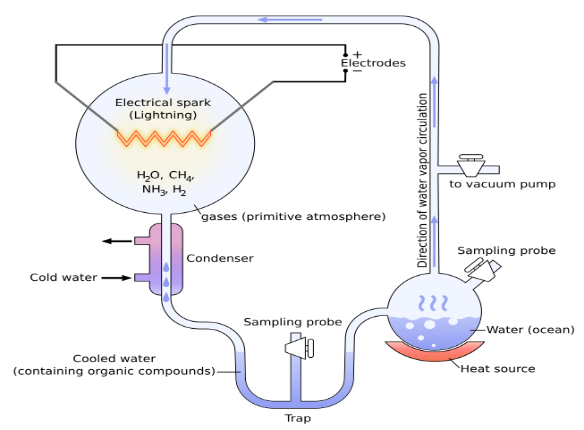
그림 3.1. 밀러-우레이 실험 장치
이 실험에 사용된 가열 장치는 수증기를 발생시키고, 진공펌프는 공기를 없애며, 방전 장치는 번개를 모방하고, 냉각 장치는 생성물을 급히 냉각시키는 장치들이다. 밀러-우레이의 실험은 거의 모든 생물학 책에서 비판 없이 진화론의 기초로 수용되고 있는데 이 실험이 모두에게 인정받기 위해서는 몇 가지 중요한 검증을 거쳐야 한다고 본다. 즉, 그들의 실험이 지구의 원시 대기를 정확히 재현하였는가 하는 검증이다. 그러기 위해서 그들이 사용한 실험 장치와 실제 지구의 원시 대기를 비교해 보도록 하자.
밀러-우레이가 자연에 있는 번개를 모방하기 위해 사용한 방전 장치는 약 5만 볼트의 전압을 사용하고 약 250도의 열을 발생시킨다. 그에 비해 번개는 약 1억 볼트 정도의 전압을 가지며 약 5만 도 정도의 열을 발생한다. 이 비교에서 밀러-우레이의 방전 장치 온도는 번개에 의해 발생하는 온도와 현저한 차이가 있음을 나타낸다. 또한 그들의 방전 장치는 방전이 주기적으로 발생하며 실험자는 혼합기체에서 화학반응이 일어나도록 방전의 빈도와 지속 시간을 인위적으로 제어할 수 있다. 이에 반해 번개는 불규칙적이며 지속시간이 수 마이크로초에서 밀리초로 상대적으로 아주 짧다. 이 비교에서 밀러-우레이의 방전 장치와 자연계의 번개는 기본적으로 아주 다름을 알 수가 있다.
지구 원시 대기를 모방하기 위해 그들이 사용한 혼합기체는 어떤가? 지구 원시 대기의 구성 성분은 혜성의 구성 성분을 분석하면 알 수 있다. 왜냐하면 혜성은 태양계를 만들고 남은 물질이기 때문이다. 아래 표에 나타나 있듯이 혜성의 구성 요소를 분석한 논문을 찾아보면 가장 많은 부분을 차지하는 것은 물(86%)이며 그다음은 이산화탄소(10%), 일산화탄소(2.6%)이고 암모니아와 메탄은 각각 1% 미만으로 아주 적다. 그런데 밀러-우레이가 사용한 혼합기체는 혜성에서 두 번째, 세 번째로 많은 이산화탄소와 일산화탄소가 빠져 있다. 이 결과는 그들이 사용한 혼합기체가 원시 대기의 구성 성분과 판이한 것을 알 수 있다.
구성 | 비율(%) | 참고문헌 |
물(H2O) | 100(86%) | Pinto et al. (2022) |
이산화탄소(CO2) | 12(10%) | Pinto et al. (2022) |
일산화탄소(CO) | 3(2.6%) | Pinto et al. (2022) |
암모니아(NH3) | 0.8(0.7%) | Russo et al. (2016) |
메탄(CH4) | 0.7(0.6%) | Mumma et al. (1996) |
표 3.1. 혜성의 구성(물=100)
밀러-우레이 실험의 다른 문제는 지구의 원시 대기가 환원성 대기였다는 가정이다. 여기서 환원성 대기란 산소가 대기에 존재하지 않음을 의미하고 있다. 만일 원시 대기가 산화성이었다면 아미노산은 생성되자마자 바로 산화되어 분해되고 만다. 원시 대기가 환원성이냐 아니면 산화성이냐에 따라 생명체의 가장 기본 물질인 아미노산의 생성이 좌우되기 때문에 원시 대기에 대해 많은 연구가 진행되어 왔다.
지금까지 알려진 바를 살펴보면 Miller(1953)와 Chyba & Sagan(1997)은 환원성 대기, Albeson(1966), Pinto et al. (1980), Zahnle(1986) 등은 약 환원성 대기, 그리고 Trail et al. (2011)은 산화성 대기임을 주장한다. 여기서 주목할 만한 논문은 2011년 네이처에 발표된 Trail et al.의 논문인데 그들은 명왕누대(Hadean)에 형성된 지르콘 산화량이 현재 지구상에서 발견되는 마그마의 산화량과 같음을 발견하였다. 또한 지르콘의 산화량으로 미루어 볼 때 지구 원시 대기가 태초에 환원성이었다고 하더라도 지구 탄생 약 2억 년 후부터는 더 이상 환원성 대기가 아니었을 거라고 발표하였다. 지금까지 발견된 최초의 원시세포 화석의 시기는 지구 탄생 후 약 5억 년이므로 생명체가 발생할 당시에는 이미 상당량의 산소가 지구 대기에 존재하였음을 예시한다. 또한 원시 대기에서 가장 높은 비율을 차지하는 이산화탄소는 산화성 기체이다. 이 사실은 그 당시 대기가 산화성임을 제시하며 아미노산이 자연적으로 생성될 가능성이 희박하였음을 의미한다.
밀러-우레이 실험의 또 다른 핵심 문제는 아미노산의 키랄성(chirality)이다. 실험실에서 생성된 아미노산은 L형과 D형이 약 50%씩 존재한다. 따라서 아미노산이 자연적으로 생성되었다면 L형과 D형의 아미노산이 약 50%씩 구성되어야 한다. 하지만 지구상의 생명체를 구성하는 아미노산은 거의 모두가(99.7%) L형 아미노산으로 구성되어 있으며 D형 아미노산은 약 0.3%밖에 되지 않는다.
현대 화학과 생명공학에서는 완벽하게 통제된 실험실 환경에서 순도 높은 출발 물질, 정교한 촉매, 최적화된 온도·압력·pH 조건을 활용하면 생명체를 구성하는 20종의 L-형 아미노산을 인공적으로 합성할 수 있다. 그러나 이러한 과정은 고도로 설계된 조건 아래 특정 목적을 위해 이루어지는 ‘인위적 합성’일 뿐, 원시 지구의 가혹하고 비선택적 환경에서 무작위로 일어나는 자연발생 과정과는 본질적으로 다르다.
게다가 지금까지 원시 지구 조건을 재현한 어떤 실험에서도 20종의 L-형 아미노산이 동시에 생성된 사례는 단 한 번도 보고된 적이 없다. 더 나아가, 설령 모든 아미노산이 우연히 생성되었다고 가정하더라도, 이들이 정확한 서열로 중합되어 74기능적 단백질을 형성하거나, 혹은 RNA와 DNA 같은 핵산을 스스로 만들어내는 것은 확률적으로 거의 불가능하다.
그렇다면 이제 먼저, 생명 기원의 핵심 열쇠로 제시되는 RNA가 과연 원시 지구 환경에서 자연적으로 생성될 가능성이 실현 가능한 수준인지, 아니면 사실상 불가능에 가까운지를 자세히 살펴보도록 하자.
ii. RNA의 형성
RNA의 자연적 형성이 얼마나 어려운지를 구체적으로 알아보려면 RNA가 어떤 구성 요소로 이루어져 있는지 알아야 한다.
RNA는 수십~수천 개의 뉴클레오타이드(nucleotide)가 인산디에스테르 결합으로 연결된 고분자이다. 하나의 뉴클레오타이드는 다음 세 부분으로 이루어져 있다: 질소 염기 (아데닌 A, 구아닌 G, 사이토신 C, 우라실 U), 오탄당인 리보스(ribose), 그리고 1~3개의 인산기(phosphate group)이다.
따라서 RNA 한 분자가 자연적으로 생기려면 이 세 구성 요소(질소 염기 + 리보스 + 인산기)가 먼저 원시 지구 환경에서 자연적으로 생성·정제된 뒤, 탈수축합 반응을 통해 정확한 순서로 수백~수천 번 결합되어야 한다.
그러면 과연 이 세 가지 구성 요소가 원시 지구 환경에서 무작위로 생성될 수 있는지, 그리고 그 확률은 어느 정도인지 하나씩 살펴보도록 하자.
- 질소염기 형성
질소염기는 고리구조를 가진 복잡한 분자들이다. 고리구조를 형성하기 위해서는 특정한 화학 반응, 특정한 반응 조건, 그리고 반응 촉매가 필요하기 때문에 단순한 원시 화합물로부터 자연적으로 생성되기 어렵다. 특정한 화학반응은 탄소 골격에 아민기(NH2 )가 첨가되는 아민화 반응, 산소를 제거하는 탈산소 반응, 고리 구조 형성 반응, 질소염기 첨가 반응 등이 있다. 반응 조건과 촉매의 경우 아민화 반응에서는 암모니아와 같은 질소 화합물과 알데히드 또는 케톤이 필요하며 반응을 촉진하는 촉매나 고온의 환경이 필요하다. 탈산소 반응에서는 환원제가 필요하며 수소나 메탄 같은 환원성 기체가 필요하다. 고리 형성 반응에서는 다단계의 고리 형성이 필요하며 금속 이온 같은 특정한 촉매, 고온 고압의 조건, 최적의 pH가 필요하다. 질소염기 첨가 반응은 다양한 전구체 화합물과 함께 고에너지 조건이 필요하다. 원시 지구 대기에서 이러한 반응들이 자연적으로 일어나려면 적절한 온도, 압력, pH, 전구체 화합물의 충분한 농도, 촉매의 존재 등 여러 조건이 정확하게 맞아야 한다. 하지만 변화무쌍한 원시 지구 대기에서 이러한 조건이 우연히 갖추어질 가능성은 극히 낮다. 실제로 실험실에서 인위적으로 최적의 환경을 조성해도 질소염기의 생성 수율은 매우 낮은 편이다. 게다가 설령 자연 상태에서 질소염기가 형성되었다고 해도, UV, 방사선, 가수분해, 산화 반응 등의 영향으로 쉽게 분해되어 축적되기 어려웠을 것이다.
- 리보스 당의 형성
오탄당인 리보스는 포르모오스 반응(formose reaction)으로 생성되는데 포름알데히드에서 시작하여 글리콜알데히드, 글리세르알데히드, 에리트로스를 생성한 후 최종적으로 리보스가 생성된다. 포르모오스 반응은 충분한 농도의 포름알데히드를 필요로 하는데 원시 대기 조건에서 포름알데히드는 쉽게 중합되거나 다른 화합물과 반응할 수 있어 안정적인 생성이 불가능하다. 또한, 포르모오스 반응에 의해 생성되는 리보스는 수율이 낮으며 포르모오스 반응의 촉진을 위해 칼슘 수산화물 같은 염기성 촉매가 필요한데 이 촉매가 존재하지 않았을 수도 있고 존재하였더라도 원시 대기 상태에서 이 촉매가 안정적이지 않고 분해될 수도 있었을 것이다. 또한 효율적인 포르모오스 반응을 위해 pH가 7 이상의 알칼리성 조건이 필요하며 적절히 높은 30~100°C의 온도가 필요하다.
이런 어려움을 극복하고 리보스가 생성되었다 하더라도 RNA 합성에 유용하려면 선택적으로 합성되고 안정화되어야 한다. 그러나 포르모오스 반응은 리보스의 선택적 생성을 선호하지 않으며, 결과적으로 여러 복잡한 당 혼합물을 생성하여 리보스만을 선택하여 RNA 합성에 활용하기 어렵게 만든다. 리보스를 복잡한 혼합물에서 선택하는 메커니즘으로 붕산염 광물과 같은 잠재적인 안정화제가 제안되었지만, 원시 대기 조건에서의 그 가용성과 효능성은 검증되지 않았다. 또 다른 심각한 문제는 리보스의 안정성이다. 리보스는 오탄당인 당 성분이라 화학적으로 불안정하여 생성되어도 수용액에서 쉽게 가수분해되거나 마이야르 반응이나 캐러멜화 같은 과정을 통해 분해되고 만다.
- 인산기의 생성
원시대기에서 인산기가 어떻게 생성되었는지 잘 알려져 있지 않다. 왜냐하면 인산 자체의 생성이 쉽지 않기 때문이다. 인산은 일반적으로 인회석(apatite)과 같은 광물에서 발견되는데, 물에 잘 용해되지 않아 원시 대기와 같은 수성 환경에서 사용 가능한 인산이 상대적으로 희귀했다. 또한 화학적 반응성이 낮아 인산기 생성에 큰 장벽을 제공한다. 뉴클레오타이드 합성에 필수적인 인산 에스터의 생성에는 상당한 에너지와 촉매 과정이 필요한데 원시 지구에서 이런 조건이 갖추어지기 쉽지 않았을 것으로 추정된다. 인산기의 사슬인 폴리인산의 생성은 높은 온도와 특별한 촉매가 필요하며 생성이 되었더라도 가수분해에 의해 단순한 인산 화합물로 분해되기 쉽다. 일부 실험실 조건에서 인위적으로 인산 분자의 생성이 가능함을 보여 주었지만 초기 지구 환경을 현실적으로 반영하지 않은 고도로 통제된 조건에서 행하여진 실험이었다. 하지만 그런 통제된 실험실 조건 아래에서도 인산염 분자의 생성 수율은 낮기 때문에 원시 대기 상태에서 생명의 기원을 유도하기 위한 충분한 양의 인산 생성은 어렵다는 것을 알 수가 있다.
- 기능성 RNA가 생성될 확률
앞에서 살펴본 대로 RNA 뉴클레오타이드를 구성하는 3가지 구성원들이 원시 지구 대기에서 자연적으로 발생할 확률은 희박함을 보았다. 하지만 모든 난관을 극복하고 질소염기, 리보스, 인산기가 성공적으로 생성되었다고 하더라도 RNA 본연의 기능을 하는 기능성 RNA의 생성이라는 또 다른 중요한 문제에 직면하게 된다. RNA에는 여러 종류가 있다. 단백질 합성에 관여하는 RNA(mRNA, rRNA, tRNA 등), 전사 후 변형에 관여하는 RNA(snRNA, snoRNA 등), 조절/규제 RNA(aRNA, miRNA 등), 그리고 기생 RNA가 있다. RNA 분자의 뉴클레오타이드 수는 그 종류에 따라 다른데 몇 가지 예를 들면 다음과 같다:
- mRNA 및 rRNA – 수백에서 수천 개
- tRNA – 70에서 90개
- snRNA – 100에서 300개
- miRNA – 20에서 25개.
여러 역할을 수행하는 RNA 분자가 평균 100개의 뉴클레오타이드로 구성되어 있다고 가정하자. 그러면 RNA 서열의 각 위치는 아데닌, 유라실, 사이토신 또는 구아닌 중 하나로 채워질 수 있는데, 길이가 100인 뉴클레오타이드 서열의 가능한 총수는 4100(=1.6x1060)이고 그중 기능성 RNA가 생성될 확률은 1/1.6x1060=6.2x10-61이 된다. 이 확률은 전 우주적 시간이 흘러도 결코 일어날 수 없는 확률이며, 이렇게 극도로 낮은 확률은 질소염기, 리보스, 그리고 인산기가 이미 존재하였더라도 기능성 RNA가 자발적으로 생성될 수 없음을 시사한다.
iii. 단백질의 형성
단백질은 아미노산의 중합에 의해 펩타이드가 형성되고 특정 3차원 형태로 적절히 접혀 있어야 제 기능을 하는 단백질이 된다.
아미노산에서 펩타이드가 형성되는 과정은 두 아미노산이 반응하여 한 아미노산의 카복실기와 다른 아미노산의 아미노기가 펩타이드 결합을 이루는 것으로 시작한다. 이 과정에서 생성되는 물 분자는 축합 반응을 통해 방출된다. 이렇게 형성된 분자는 펩타이드 결합을 통해 연결되며 이 과정이 반복되어 긴 펩타이드 사슬이 형성된다. 이 사슬을 단백질의 1차 구조라 부른다.
1차 구조의 폴리펩타이드 사슬은 알파 나선 및 베타 시트로 알려진 2차 구조로 접힌다. 그리고, 이러한 구조는 폴리펩타이드 사슬의 백본 원자들 간의 수소 결합에 의해 안정화된다. 나선과 시트를 연결하는 추가적인 2차 구조, 턴과 루프는 단백질의 전체 접힘에 기여를 한다. 2차 구조는 더 나아가 소수성 상호작용, 수소 결합, 이온 결합, 및 이황화 결합에 의해 특정한 3차원 형태(3차 구조)로 접힌다. 소수성 상호작용에서는 비극성 사슬이 수성 환경에서 벗어나 클러스터링 되어 폴리펩타이드가 압축된 구형 형태로 접히게 된다. 수소 결합은 극성 사슬과 백본 간에 형성되어 접힌 구조를 안정화한다. 이온 결합은 반대 전하를 가진 사슬 간의 정전기적 상호작용으로 단백질의 안정성에 기여하며, 이황화 결합은 시스테인 잔기 간의 공유 결합으로 추가적인 안정성을 제공한다.
여러 폴리펩타이드 사슬(소단위체)로 구성된 일부 단백질의 경우, 이러한 접힌 단위들이 함께 모여 4차 구조를 형성한다. 오류를 방지하기 위해 샤페론 단백질이 접히는 과정을 도와 잘못 접히거나 뭉쳐지는 것을 방지하여 올바른 형태를 취하도록 돕는다. 단백질은 가장 안정적이고 기능적인 형태를 얻기 위해 약간의 형태 변화를 하거나 인산화, 당화, 절단 등 화학적 수정을 하기도 한다.
아미노산에서 단백질이 만들어지는 여러 과정은 아주 복잡하지만 정교하게 잘 짜인 알고리즘에 따라 진행되는 것을 알 수가 있다. 이 사실과 단백질이 만들어지는 기본 설계도가 이미 DNA에 이미 들어 있다는 사실도 단백질의 형성이 방향성 없는 무작위 과정이 아닌 정교한 지적 설계의 산물임을 시사한다.
그림 3.2. 단백질 형성 과정
확률적으로 보아도 무작위 과정으로 단백질이 형성되기는 어렵다. 하나의 단백질에 포함된 아미노산의 수는 수십에서 수천 개가 되는데, 소형 단백질인 인슐린에는 약 51개, 중형 단백질인 미오글로빈에는 약 153개, 대형 단백질인 헤모글로빈에는 약 574개, 거대 단백질인 타이틴에는 약 34,350개의 아미노산 조합이 들어있다. 20종류의 아미노산 조합에 의해 소형 단백질인 인슐린이 형성될 확률은 1/2051=4.4x10-67이며 거대 단백질인 타이틴이 형성될 확률은 1/2034350=10-44680≈0이다. 이 역시 단백질 형성이 무작위 과정으로는 형성되기는 불가능하다는 것을 시사한다.
아미노산에서 펩타이드 결합이 되는 과정은 많은 에너지를 필요로 하는데, 원시 대기 상태에서 이러한 결합을 촉진할 일관되고 충분한 에너지원이 존재하였는지에 대한 의문도 제시된다. 번개, 자외선, 방사선, 화산 열과 같은 다양한 에너지원이 제안되었지만, 이러한 에너지원이 일관되게 펩타이드 결합 형성을 촉진할 수 있는 효율성과 신뢰성은 논란의 여지가 많다. 왜냐하면 원시 지구의 환경은 극한의 온도, pH 수준 및 환경 변화와 함께 가혹하고 변동이 심했을 가능성이 크기 때문이다. 이러한 조건은 오히려 펩타이드 결합 형성의 섬세한 과정을 방해하고, 형성된 펩타이드의 안정성을 저해할 수 있다. 또한 펩타이드는 수용성 환경에서 가수분해되기 쉬워 펩타이드가 형성되기보다는 빠르게 분해될 수도 있다.
iv. DNA의 형성
DNA 뉴클레오타이드는 RNA와 마찬가지로 세 부분으로 구성된다: 질소염기(구아닌, 아데닌, 사이토신, 티민), 디옥시리보스 당, 그리고 인산기다. 이들의 자발적 형성은 RNA에서 보았듯이 거의 불가능하다. DNA의 자발적인 형성에 있어 또 다른 난관은 RNA와 달리 추가적으로 이중 나선 구조의 염기쌍을 형성하여야 한다는 것이다. DNA 이중 나선은 아데닌과 티민, 사이토신과 구아닌이 특정한 순서로 배열되어 있으며 반대 가닥에는 안전성을 위해 보완적인 서열이 존재한다. 이렇게 두 개의 보완적인 서열이 완벽하게 정렬된 구조이기 때문에 안내 템플릿이나 메커니즘 없이 무작위적으로 만들어지는 것은 불가능하다.
자가복제는 DNA의 주요 기능 중 하나이다. DNA 복제는 정확성과 신뢰성을 보장하기 위해 복잡한 효소와 단백질이 필요하다. DNA 복제에 관여하는 주요 효소는 헬리케이스, 단일 가닥 결합(SSB) 단백질, 프라이메이스, DNA 폴리메라아제, 리보뉴클레아제 H(RNase H), DNA 리가아제, 토포아이소머라아제 등이 있다. DNA 복제에 참여하는 효소의 아미노산 총수는 대개 수백 개에서 수천 개사이다. DNA 복제에 이런 효소들이 없으면 이중 나선 복제 및 오류 수정이 되지 않는다. 그런데, 이 효소 중 어느 하나라도 우연히 생성될 확률은 사실상 0이다. 예를 들어, RNase H가 우연히 생성될 확률은 20-155≈2.2x10-202≈0으로 현실적으로 발생할 확률이 전혀 없다.
DNA는 긴 단일 가닥으로 있는 게 아니라 염색체 속에서 매우 복잡하게 접힌 뉴클레오솜 형태로 존재한다. DNA가 접히는 과정을 DNA 응축 과정(DNA condensation process)이라고 하는데 이 과정은 많은 단백질의 도움으로 여러 단계의 압축과 접힘 과정을 거쳐 이루어진다. 이중 나선 구조를 가진 DNA는 히스톤 단백질과 결합하여 뉴클레오솜을 형성한다. 뉴클레오솜은 히스톤 단백질 코어(옥타머: H2A, H2B, H3, H4 각각 2개씩)와 이를 약 1.65회 감싼 147개의 염기쌍으로 구성된 DNA로 이루어져 있다. 그 구조는 마치 구슬(뉴클레오솜)이 줄(링크 DNA)에 매달려 있는 것처럼 연결되어 있다. 이 뉴클레오솜들은 추가로 감겨 30nm 직경의 섬유 모양을 형성하여 나선형으로 배열되거나 지그재그 형태로 배열되어 있다. 30nm 섬유 구조는 루프 도메인이라는 더 큰 구조로 접힌다. 이 루프 도메인은 특정 단백질에 의해 핵 내 특정 위치에 고정되어 있으며, 유전자 발현 및 기타 기능적 활동을 조절하는 역할을 한다. 루프 도메인은 더욱 복잡하게 접히고 압축되어 염색체를 형성하며, 염색체는 세포 분열 시 DNA를 딸세포에 정확하게 전달하기 위해 가장 높은 수준으로 압축되어 있다. 이 단계에서 코헤신과 콘덴신 같은 단백질 복합체가 염색체 구조를 안정화하고, 올바르게 분리되도록 돕는다.
이상에서 살펴본 바와 같이, DNA의 형성과 복제 과정은 매우 복잡하며 다양한 생화학적 법칙과 여러 효소의 정교한 협력이 필수적으로 요구된다. 이렇게 복잡한 과정들에 대한 진화론적 설명은 명확하지 않으며 단순히 DNA가 RNA에서 전환되었다고만 주장한다. 하지만, 이 주장이 성립하기 위해서는 RNA가 어떻게 형성되었는지, RNA에서 DNA의 이중 나선 구조가 어떤 메커니즘으로 형성되었는지, 그리고 DNA 복제에 필수적인 다양한 효소들이 어떤 과정으로 생겨났는지 등을 구체적으로 설명해야 한다. 이러한 점을 고려할 때, DNA는 단순한 무작위적 과정의 산물이 아니라 정교하게 계획된 지적 설계의 산물임을 강하게 시사한다.
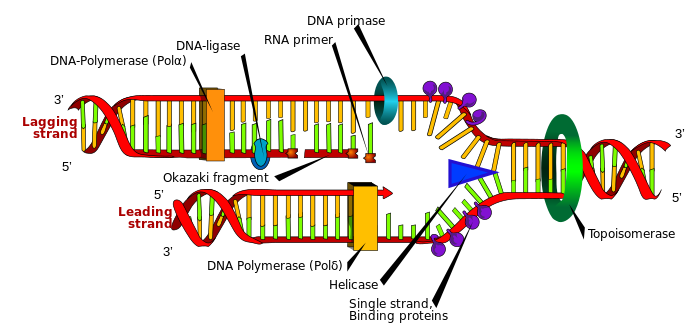
그림 3.3. DNA 복제 과정
v. 원핵세포의 형성
논의를 계속하기 위해 RNA, 단백질, DNA가 우연히 형성되었다고 가정하면 생명체를 향해 그다음 넘어야 하는 산은 세포의 형성이다. 세포에는 두 가지 주요 유형이 있다: 원핵세포와 진핵세포다. 박테리아와 고세균과 같은 생물에서 발견되는 원핵세포는 단순하며 핵이 없다. 유전 물질은 세포질에 자유롭게 떠다니는 단일 원형 DNA 분자에 포함되어 있으며 세포소기관들도 없다. 과학자들은 원시세포가 자연 선택, 돌연변이 및 환경 적응 등 점진적인 과정을 통해 원핵세포로 진화했다고 주장한다. 하지만 원시세포의 존재는 확인된 바가 없으며 원핵세포가 지구상에 처음 등장한 세포였을 것으로 간주한다. 화석 기록에 따르면 원핵세포는 약 35억 년에서 38억 년 전에 지구에 나타났다. 모든 세포는 세포막으로 둘러싸여 있기 때문에 원시 대기에서 세포가 자연스럽게 형성되었다면 그 첫 번째 과정은 이 세포막의 형성일 것으로 추측된다. 그렇다면, 세포막이 무작위적 과정을 거쳐 자연적으로 형성될 수 있는지 알아보자.
- 세포막 형성
세포막은 단순한 구조로 생각되지만 실제로는 지질(인지질, 콜레스테롤, 당지질), 단백질, 그리고 탄수화물로 구성된 복잡하고 역동적인 구조이다. 인지질은 세포막의 이중층 구조를 형성하고, 콜레스테롤은 유동성을 조절하며, 당지질은 세포 인식에 기여한다. 단백질은 내재성 단백질과 주변부 단백질로 나뉘며, 운반, 신호 전달, 구조적 지지 등의 역할을 한다. 탄수화물은 세포 인식과 통신에 중요한 역할을 한다.
원시 지구에서 기능성 막(functional membrane)에 필요한 특정 양친성(amphiphilic) 지질 분자가 형성되기 위해서는 지방산, 글리세롤, 그리고 인산기가 필요하다. 하지만, 역동적인 원시 지구의 환경에서는 자발적으로 이런 분자들의 정확한 조합이 형성되기 어렵다. 더구나, 앞에서 살펴보았듯이 인산기의 자발적인 형성은 불가능하며 이 인산기가 없이는 세포막이 형성될 수가 없다.
하지만, 어떻게 하여 양친성 지질 분자가 형성되었다 하더라도 세포 환경을 캡슐화하고 보호할 수 있는 안정적이고 반투과성의 이중층이 형성되어야 한다. 그런데, 이 이중층을 형성하기 위해서는 특정한 조건이 필요한데, 충분한 농도의 양친성 분자(예: 인지질), 중성에 가까운 pH, 막의 안전성을 높여 줄 칼슘이나 마그네슘과 같은 이온의 존재, 높거나 낮은 온도가 아닌 적절한 온도, 이중층 형성에 필요한 수용성 환경, 자외선, 열, 전기적 방전 등과 같은 에너지원, 그리고 촉매 표면의 존재등 이다. 하지만, 초기 지구의 조건은 극한의 온도, 변동스러운 pH 상태, 강한 방사선 등으로 가혹하고 변동이 심하였으며, 이러한 환경에서 이중층 막이 형성되었다 하더라도 안정성을 유지하기 어려웠고 쉽게 파괴되었을 것이다.
만일, 원핵세포의 크기를 박테리아 세포와 같은 1마이크로미터 정도로 가정하면, 그 표면적은 3x10-12m2가 된다. 단일 인지질 분자의 크기는 약 5x10-19m2이므로 이중층을 형성하기 위해 필요한 인지질의 수는 약 1,000만 개가 된다. 이 1,000만 개의 인지질이 나란히 정렬되는 것도 힘들지만 어떤 형태의 안내나 방향성 없이 스스로 동그랗게 굽어져 3차원의 폐쇄된 세포막을 만들었을 가능성은 거의 없을 것이다.
세포막은 필수 영양소와 분자를 선택적으로 통과시키면서 유해 물질을 차단하는 기능을 가져야 한다. 이러한 선택적 투과성은 복잡한 단백질과 채널의 존재를 필요로 하는데 이들이 무작위 과정으로 형성되고 스스로 알아서 세포막에 통합될 가능성은 극도로 낮다. 이렇게 세포 형성의 첫째 단계인 세포막이 우연히 형성될 확률이 희박하다면 생명체를 향한 그다음 단계로는 나아갈 수가 없다. 또한, 세포막이 어떻게 하여 자연적으로 형성되었다 하더라도, 세포막 내부를 구성하는 세포질, 리보솜, 플라스미드 등이 어떻게 형성되었는지에 대한 진화론적 설명이 부족하다. 그리고 이들이 어떻게 하여 우연히 형성되었더라도 어떤 메커니즘을 통해 세포막 속에 캡슐화될 수 있었는지에 대한 추가적인 설명도 필요하다.
vi. 진핵세포의 형성
진핵세포 기원에 대해 널리 받아들여지는 이론은 세포내 공생설(endosymbiotic theory)이다. 공생설은 진핵세포가 원시 원핵세포 간의 공생 관계를 통해 기원했다고 제시한다. 이 과정은 특정 원핵세포(동물 세포의 경우 미토콘드리아, 식물 세포의 경우 엽록체)가 조상 숙주세포(ancestral host cell)에 의해 흡수되어 상호 유익한 관계를 형성하고 결국 복잡한 진핵세포로 발전하였다고 한다. 그럴듯한 시나리오로 들리지만, 이 이론이 성립하기 위해서는 조상 숙주세포와 원핵세포가 이미 존재하고 있어야 한다. 조상 숙주세포로는 고세균(archaea)을 주장하는데 문제점은 고세균에서 원핵세포를 세포 안으로 끌어들이는 내포작용(endocytosis)이 전혀 관찰되지 않았다는 점, 그리고 고세균의 세포막은 에테르 결합으로 구성됐지만 진핵세포 세포막은 에스테르 결합으로 구성되어 구조적으로 큰 차이를 보인다는 점이다.
앞서 원핵세포가 자연적으로 생성되기 어렵다고 했는데, 같은 원핵세포에 해당하는 미토콘드리아 역시 자연적으로 생성되기는 어려울 것으로 예상된다. 미토콘드리아는 산화적 인산화를 통해 ATP를 생성하는 세포의 발전소 역할을 하는 복잡한 세포소기관이다. 미토콘드리아는 외막, 막간 공간, 내막 및 기질(효소, DNA, 리보솜, 대사 산물) 등으로 구성되어 있다. 세포막과 마찬가지로 외막은 인지질과 단백질의 혼합물로 이루어진 인지질 이중층으로 이루어져 있다. 우리는 앞에서 세포막이 무작위 과정으로 형성되기 어렵다고 밝혔다. 또한 외막보다 더 복잡한 DNA의 자연 형성도 불가능하기에 DNA를 포함하고 있는 미토콘드리아가 자연적으로 형성될 가능성은 희박하다.
진핵세포에서 세포핵의 기원은 더욱더 설명이 불가능하다. 세포핵은 유전 물질을 보호하고, 세포의 대사 과정을 효율적으로 조절하는 가장 중요한 세포소기관이다. 세포핵은 핵막에 쌓여 있다. 진화론에서는 세포 핵막의 기원에 대해 막 내포 가설, 바이러스 기원 가설, 유전자 전이 가설 등을 제시한다. 막 내포 가설은 원핵세포의 세포막이 안쪽으로 접혀 들어가면서 핵막을 형성했다는 가설이고, 바이러스 기원 가설은 바이러스가 원시 진핵세포와 상호작용을 하면서 유전 물질을 보호하는 핵막 구조를 형성하였다는 가설이며, 유전자 전이 가설은 서로 다른 원핵생물 간의 유전자 전이 및 혼합이 발생하면서 복잡한 유전체가 형성되고, 이를 보호하기 위한 세포막이 형성되었다는 가설이다. 하지만 이런 가설들은 다음과 같은 이유로 타당성이 없다. 막 내포 가설의 경우 세포막은 단일 인지질 이중층으로 구성되어 있지만 핵막은 두 개의 인지질 이중층(외막과 내막)으로 구성되어 있고, 핵막에는 핵공 복합체란 특수한 구조가 있지만 세포막에는 없고, 세포막과 핵막에 존재하는 단백질의 종류와 배치가 다르기 때문이다. 바이러스 기원 가설의 경우 대개 바이러스는 세포의 유전 물질을 자신의 복제를 의한 숙주 세포로 이용하는데 오히려 보호하기 위해 핵막을 형성하였다는 설명은 설득력이 없다. 유전자 전이 가설은 전이 자체가 세포 내의 유전자 다양성을 증가시킬 수 있지만 핵막과 같은 복잡한 구조의 형성을 설명하기에는 불충분하고 그 메커니즘도 불분명하다.
핵막 안에는 핵소체와 염색체가 존재하는데 이들의 기원에 대해서도 납득할 만한 설명이 없다. 핵소체와 염색체에는 생명체의 유전 정보가 들어 있으며 RNA, 단백질, DNA, 세포소기관, 생명체의 조직 및 기관을 형성하는 설계도가 들어 있다. 생명체가 형성되기 전의 진핵세포 단계에서 생명체를 구성하는 설계도가 이미 핵 속에 들어 있다는 사실은 진화론적으로는 절대적으로 설명될 수 없으며, 오히려 생명체의 지적 설계를 나타내는 명확한 증거가 된다.
공생설이 지지를 받으려면 원핵세포와 진핵세포 간의 전이 형태를 보여 주는 화석적 증거가 필요한데 이런 화석은 발견되지 않았다. 또한, 미토콘드리아는 자체 DNA와 독립적으로 복제할 수 있는 능력 등 일부 자율성을 유지한다. 만일 미토콘드리아가 진핵세포로 통합이 되었다면 수십억 년의 진화 후에도 미토콘드리아의 유전체가 숙주 유전체에 완전히 통합되지 않은 의문도 제기된다. 이에 더해 미토콘드리아를 구성하는 DNA, 효소, 리보솜 등도 자연적으로 형성되기 불가능하다.
vii. 세포소기관의 국재화
세포 내에서는 여러 세포소기관이 존재한다. 세포소기관 국재화(organelle localization)는 세포 내에서 세포소기관들이 특정 위치에 배치되는 과정이다. 핵은 세포 중앙, 미토콘드리아는 세포 주변부, 소포체는 핵 근처, 골지체는 소포체 근처, 그리고 리소좀은 세포질에 위치한다. 세포소기관들은 적절한 위치에 있을 때 세포의 기능이 최적화되고 세포의 구조적 통합성과 기능적 조화가 잘 유지된다. 세포소기관들은 스스로 생각을 할 수 없을 텐데 어떻게 하여 그런 최적의 장소로 알아서 찾아가는 것일까? 세포 소기관의 국재화 과정을 자세히 살펴보면, 그것이 매우 복잡하다는 것을 알 수 있으며, 이러한 메커니즘이 우연히 형성된 것이 아니라 정교한 설계에 의해 조율되고 진행된 과정임을 확인할 수 있다.
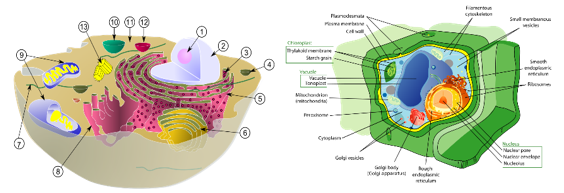
그림 3.4. 동물 세포와 식물 세포의 구조
세포소기관 국재화는 세포 골격, 모터 단백질, 막 수송, 앵커 단백질 및 스캐폴드 단백질, 동적 조정, 세포 간 통신 등의 복잡한 상호 작용으로 이루어진다.
세포 골격은 세포의 구조적 지지력을 제공하며 미세소관, 액틴 필라멘트, 중간 필라멘트의 세 가지 주요 유형의 필라멘트로 구성되어 있다. 각 필라멘트는 세포소기관 국재화의 특정한 역할을 담당한다.
미세소관은 튜불린 단백질로 이루어진 길고 속이 빈 관이며, 미세소관형성중심(중심체)에서 세포 주변으로 이어지는 네트워크를 형성한다. 미세소관은 중심체 근처에 위치한 골지체와 세포 여러 곳에 분포하는 미토콘드리아와 같은 소기관들을 키네신과 다이네인과 같은 모터 단백질의 도움으로 세포 내 적절한 곳으로 이동시키는 도로망 역할을 한다. 키네신은 세포소기관을 미세소관의 플러스 끝, 일반적으로 세포 주변으로 이동시키고, 다이네인은 마이너스 끝, 일반적으로 세포 중심 쪽으로 이동시킨다.
마이크로필라멘트라고도 알려진 액틴 필라멘트는 액틴 단백질로 만들어진 가늘고 유연한 섬유다. 액틴 필라멘트는 원형질막 바로 아래에 집중되어 있으며 세포질 전체에 촘촘한 네트워크를 형성한다. 액틴 필라멘트는 세포소기관과 영양분을 세포 전체에 분배하는 과정인 세포질 스트리밍 역할을 촉진한다. 미오신 모터 단백질은 액틴 필라멘트와 상호작용을 하여 액틴 네트워크를 따라 소포체, 엔도솜 및 기타 작은 소기관을 운반한다. 액틴 필라멘트는 세포 모양을 유지하고 세포 이동에 관여하여 소기관의 위치에 간접적으로 영향을 미친다.
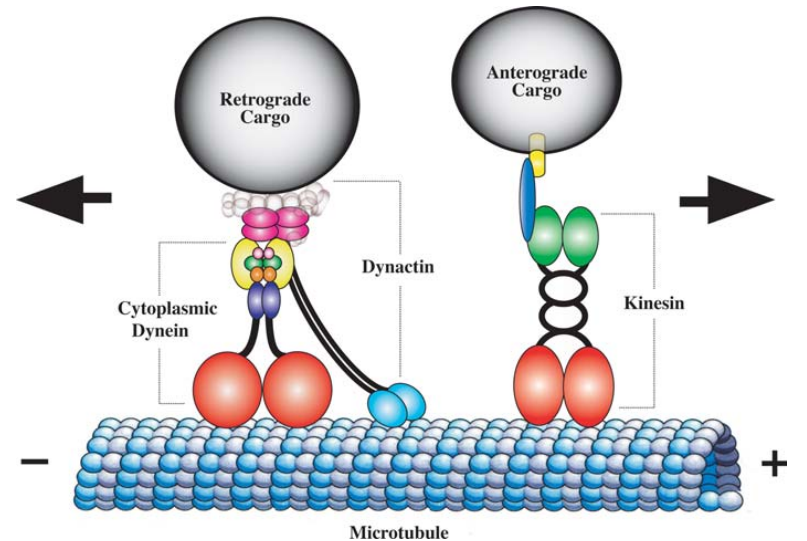
그림 3.5. 미세소관과 모터 단백질
중간 필라멘트는 세포 유형에 따라 다양한 단백질(케라틴, 비멘틴, 라민 등)로 구성된 밧줄 같은 섬유로, 세포의 기계적 강도와 구조적 지지력을 제공한다. 중간 필라멘트는 핵과 같은 소기관을 세포질 내에 제자리에 고정하여 위치를 안정화하는 데 도움을 준다. 또한 세포 골격의 전체적인 무결성을 유지하여 미세소관 및 액틴 필라멘트와 같은 다른 구성 요소가 세포소기관의 국재화에 효과적으로 기능할 수 있도록 한다.
다양한 유형의 세포 골격 필라멘트는 종종 함께 작용하여 세포소기관을 정확하게 배치한다. 예를 들어, 미세소관과 액틴 필라멘트는 소포(vesicle)와 소기관의 적절한 분포와 이동을 보장하기 위해 협력한다. 세포 골격은 매우 역동적이며 세포의 필요에 따라 지속적으로 리모델링된다. 이러한 유연성 덕분에 세포 신호나 환경 변화에 따라 세포소기관을 신속하게 재배치할 수 있다.
막 수송은 세포 내에서 소포를 통해 단백질, 지질 및 기타 분자를 세포 내의 정확한 목적지에 수송하는 과정이다. 클라트린 같은 특정 단백질이 관여하여 기증되는 막의 특정 부위에서 소포가 형성되고 운반할 화물 분자를 소포로 포장한다. 형성된 소포는 모터 단백질의 도움으로 세포골격 내의 액틴 필라멘트와 미세소관을 따라 이동된다. 그렇게 하여 소포가 목적지에 도착하면 목적지의 막과 융합하여 내용물을 방출한다. 이 과정에서 SNARE 단백질과 같은 특수 단백질이 소포와 목표 지점의 막을 인식하고 결합을 돕는다. 막 수송에 관여하는 주요 세포소기관으로는 소포체, 골지체, 엔도솜 및 리소좀과 같은 다양한 유형의 소포체가 있다. 이 과정은 세포 조직을 유지하고, 소기관 간의 통신을 촉진하며, 세포가 내부 및 외부 신호에 효율적으로 반응할 수 있도록 하는 데 필수적이다.
신호 경로는 공간적 단서를 제공하는 화학적 신호 전달을 통해 세포소기관이 적절한 위치로 이동할 수 있도록 한다. 세포질 표면과 세포질 내의 수용체는 이 과정을 촉진하기 위해 신호 분자와 상호 작용을 한다. 예를 들어, Rab 단백질과 같은 작은 GTPase는 특정 이펙터 단백질과 상호작용을 하여 소포 이동과 소기관 위치를 제어하는 핵심 조절 인자이다. 이러한 신호 경로는 변화하는 세포의 요구와 환경 조건에 따라 세포 과정이 조정되고 세포소기관이 동적으로 배치되도록 한다.
앵커 단백질과 스캐폴드는 세포소기관이 세포 내에 정확하게 위치하도록 함으로써 세포 위치 결정에 중요한 역할을 한다. 앵커 단백질은 세포질 내의 특정 부위에 소기관을 연결하여 소기관을 안정화하고 소기관의 이동을 방지한다. 예를 들어, 미토콘드리아는 특정 고정 메커니즘을 통해 소포체에 고정되어 효율적인 에너지 전달과 대사 조정을 촉진할 수 있다. 스캐폴드 단백질은 세포소기관을 제자리에 고정하는 복합체를 형성하여 세포의 전체 조직을 유지함으로써 구조적 지원을 제공한다. 이러한 단백질은 세포소기관의 적절한 배열을 가능하게 하는 동적 프레임워크를 만들어 세포 기능이 효과적이고 효율적으로 수행될 수 있도록 한다.
세포 위치의 동적 조정은 세포 내 소기관의 위치가 연속적이고 반응적으로 조정 정렬되는 과정을 말한다. 이러한 조정은 세포의 기능과 적응력을 유지하는 데 매우 중요하다. 유사 분열과 같은 세포 주기의 여러 단계에서 핵과 미토콘드리아와 같은 세포소기관은 적절한 세포 분열을 위해 위치를 동적으로 변경한다. 또한 영양소 가용성이나 스트레스 조건과 같은 환경적 자극에 반응하여 세포소기관은 그 기능이 가장 필요한 영역으로 재배치될 수 있다. 이러한 동적 재배치는 세포 골격과 모터 단백질에 의해 촉진되어 세포가 항상성을 유지하고 변화하는 내부 및 외부 조건에 효율적으로 대응할 수 있도록 한다.
세포 간 통신은 세포 내 다양한 소기관들이 서로 신호를 주고받거나 물질을 교환하면서 협력하는 과정을 말한다. 세포는 이러한 통신을 통해 생리적 기능을 조절하고 항상성을 유지하며, 외부 자극에 효과적으로 대응할 수 있다. 세포 간 통신은 직접 접촉 부위와 소포체 수송을 통해 이루어지는데 세포의 기능을 조율하고 항상성을 유지하는 데 필수적이다. 미토콘드리아와 소포체 사이의 미토콘드리아 관련 막(MAMs)과 같은 접촉 부위는 지질, 칼슘 및 기타 분자의 이동을 촉진하여 세포소기관 간의 동기화된 활동을 보장한다. 소포 수송은 소포의 분화와 융합을 포함하며, 소포는 세포소기관 간에 단백질과 지질을 운반하여 세포소기관의 기능적 통합을 유지한다. 효과적인 세포 간 통신은 신진대사, 신호 전달, 스트레스 반응과 같은 과정에 필수적이다.
이상에서 살펴본 바와 같이, 세포소기관의 국재화 메커니즘은 고도로 복잡하고 정교하게 조직되어 있으며, 명확한 목적과 방향성을 가지고 있다. 진화론에서는 이러한 정교한 시스템이 무작위 돌연변이와 자연 선택을 통해 점진적으로 형성되었다고 주장한다. 이 주장이 인정받기 위해서는 먼저, 모터 단백질, 앵커 단백질, 스캐폴드 단백질과 같은 핵심 구성 요소들이 무작위 돌연변이만으로 형성될 수 있는지에 대한 구체적인 설명이 필요하다. 또한, 막 수송, 동적 조정, 세포 간 통신과 같은 필수적인 메커니즘이 어떻게 확립되었는지에 대한 실질적인 증거가 제시되어야 한다. 특히, 세포소기관 국재화 메커니즘이 단계적으로 진화했다는 주장을 뒷받침할 수 있는 중간 단계의 화석이나 생물학적 증거 역시 제시되어야 한다. 더 나아가, 세포소기관 국재화에 관여하는 단백질과 메커니즘 중 어느 하나라도 빠지면 기능을 수행할 수 없다는 점에서, 이러한 복잡한 요소들이 어떻게 완벽한 조화를 이루며 동시에 존재하게 되었는지에 대한 설명이 요구된다. 하지만 진화론에서는 이런 핵심적 사실들을 전혀 설명하지 못한다.
지금까지 알아본 세포소기관의 국재화 과정은 마치 아마존에서 물건을 주문하면 정확한 주소로 배송되는 과정과 유사하다. 아마존의 물류 운송 체계가 우연히 형성된 것이 아니라 정교한 계획에 의해 구축된 것처럼, 세포 소기관의 국재화 메커니즘 또한 무작위 과정의 결과가 아니라 정교한 설계에 의해 형성되었음을 명확히 보여준다.
viii. 세포 분화
세포 분화는 비특화 세포로부터 특정한 기능을 가진 특화 세포로 분화하는 과정을 말한다. 세포 분화는 생명체의 조직 및 기관 형성에 필수적이며, 줄기세포로부터 시작해 유전자 발현 조절, 세포 외부 신호, 세포 간 상호작용 등 다양한 메커니즘에 의해 세포 분화가 조절된다.
유전자 발현 조절은 전사 인자 단백질이 특정 유전자의 발현을 활성화하거나 억제하여 세포의 운명을 결정한다. 예를 들어 MyoD는 근육세포로, Sox2는 신경세포로 분화시킨다. 이 과정은 DNA 서열의 변화 없이 유전자 발현을 조절하는 DNA 메틸화, 히스톤 수정, 비암호화 RNA 과정이 들어가는데 이를 에피제네틱 조절 과정이라 부른다. DNA 메틸화는 DNA의 시토신 염기에 메틸기를 추가하는 과정으로, 보통 유전자 발현을 억제한다. 히스톤 단백질의 수정(아세틸화 또는 메틸화)은 염색질의 구조를 변화시켜 유전자 접근성을 조절하며, 히스톤 아세틸화는 유전자 발현을 활성화하고, 메틸화는 유전자 발현을 억제한다. 비암호화 RNA(ncRNA)는 단백질로 번역되지 않지만 유전자 발현 조절과 세포 분화 과정에서 중요한 역할을 한다. 예를 들어 miRNA는 mRNA와 결합하거나 mRNA를 분해하여 유전자 발현을 조절한다. siRNA도 mRNA와 결합하여 분해함으로 특정 유전자 발현을 억제한다. lncRNA는 다양한 메커니즘을 통해 유전자 발현을 조절할 수 있으며, 염색질 리모델링, 전사 조절, 번역 조절 등에 관여한다. snRNA와 snRNP는 스플라이싱 과정에서 인트론을 제거하고 엑손을 연결하는 역할을 하며, snoRNA는 rRNA의 메틸화를 촉진한다.
세포 외부 신호는 세포 외부에서 분비된 물질이나 인자로 다양한 형태가 존재하며, 특정 세포의 표면 수용체에 의해 인식되고 세포 내 MAPK/ERK, PI3K/Akt, Wnt/β-Catenin 경로 등의 신호 전달 경로를 통해 세포 내부로 전달된다. 외부 신호의 주요 유형에는 성장 인자, 모프젠, 호르몬, 사이토카인 등이 있다. 성장 인자는 세포의 성장, 분화, 생존을 촉진하는 단백질들이며 표피세포는 EGF, 섬유아세포는 FGF, 혈관 내피세포는 VEGF 등이 있다. 모프젠은 농도 구배에 따라 세포의 운명을 결정하는 신호 분자로 신경관의 발달을 조절하는 Shh, 뼈와 연골 형성을 유도하는 BMPs 등이 있다. 호르몬은 잘 알려진 대로 인슐린, 에스트로겐, 테스토스테론 등이 있으며 혈류를 통해 표적 세포로 이동하여 특정 생리적 반응을 유도하는 신호 분자다. 사이토카인은 면역 반응을 조절하는 단백질로 면역세포 간의 신호를 전달하는 인터류킨, 바이러스 감염에 대한 면역 반응을 촉진하는 인터페론, 염증 반응과 세포 사멸을 유도하는 TNF 등이 있다.
세포 간 상호작용은 세포가 서로 신호를 교환하여 세포 운명을 결정하고, 조직의 형성 및 기능을 조절하는 과정이다. 이러한 상호작용은 세포의 표면에 있는 수용체와 리간드 간의 결합을 통해 이루어지며, 특정 신호 전달 경로를 활성화하여 세포 분화를 유도한다. 세포 간 상호작용 메커니즘으로는 Notch 신호 전달 경로, 간극 결합(gap junctions), 카드헤린과 인테그린이 간여하는 접촉 의존적 신호 전달, 가까운 세포로 신호를 전달하는 파라크린 신호 전달, 모프젠 농도 구배를 통한 신호 전달 등이 있다.
이상에서 살펴본 대로 세포 분화 과정은 유전자 발현, 외부 신호, 세포 간 상호작용 등의 여러 복잡하고 정교한 메커니즘들이 서로 잘 협력이 되어 진행되고 있음을 알 수가 있다. 유전자 발현 조절의 경우 세포 분화의 특정 단계와 장소에서 올바른 신호를 받았을 때만 특정 유전자가 활성화된다. 이 과정에서 수백 개의 전사인자, 인핸서, 리프레서 및 기타 조절 요소들이 상호작용을 하는 복잡한 네트워크를 통해 유전자 발현을 조절한다. 세포 외부 신호는 특정 리간드가 특정 수용체에 결합하며 다단계 신호 전달 경로를 활성화시키고 세포 내부에서 특정 반응을 유도한다. 세포 간 상호작용도 특정 세포가 특정 위치에서 특정한 역할을 하도록 유도되며 양성 및 음성 피드백에 의해 조절된다.
진화론에서는 세포 분화 역시 돌연변이와 자연 선택을 통해 점진적으로 형성되었다고 주장하지만 이를 뒷받침할 구체적인 증거가 부족하다. 먼저, 세포 분화는 단순한 생화학적 반응이 아니라 명확한 목적과 방향성을 가지고 있지만 돌연변이와 자연 선택은 이런 방향성이 없다. 진화론에서는 단세포 생물이 점진적으로 다세포 생물로 발전하면서 세포 분화가 이루어졌다고 주장하지만, 단세포에서 다세포로의 전환 과정, 그리고 조직과 기관이 형성되는 중간 단계를 입증할 실질적인 화석적 및 생물학적 증거는 존재하지 않는다. 현재 발견된 다세포 생물의 화석들은 이미 완전한 형태로 분화된 상태이며, 점진적으로 복잡성이 증가하는 과정을 보여주지 않는다. 또한, 세포 분화는 유전자 발현, 외부 신호, 세포 간 상호작용이 정밀하게 조율된 상태에서 동시에 작동해야 하는 과정이며 어느 하나라도 빠지거나 아직도 진화 과정이라면 작동하지 않는다. 이런 여러 사실은 세포 분화가 무작위적인 돌연변이와 자연 선택의 산물이 아니라, 처음부터 정교하게 설계된 지적 설계의 결과임을 분명히 보여준다.
ix. 조직과 기관의 형성
생명체를 향한 세포 분화의 다음 단계는 조직과 기관의 형성이다. 세포의 조직 형성은 줄기세포가 세포 간 상호작용, 세포 이동, 세포 외 기질의 복합적인 작용에 의해 세포들이 모여 특정 기능을 수행하는 조직을 형성하는 과정이다.
세포 간 상호작용은 세포들이 서로 신호를 교환하고 물리적으로 결합하여 조직을 형성하는 과정이다. 상호작용 신호 전달은 Notch 신호 경로와 Wnt 신호 경로를 사용한다. 결합에 관여하는 접착 단백질 분자로 카드헤린은 세포 간의 결합을 매개하여 조직의 구조적 통합성을 유지하고, 인테그린은 세포와 세포 외 기질 간의 결합을 매개하여 조직의 안정성을 제공한다.
세포 이동은 세포가 특정 위치로 이동하여 조직을 형성하는 과정이다. 화학 신호를 따라 세포가 이동하며(케모택시스), 미세섬유(액틴 필라멘트) 같은 세포 골격 요소들이 재구성되어 세포 이동을 촉진한다. 세포 이동은 배아 발달, 상처 치유, 면역 반응 등에 중요한 역할을 한다.
세포 외 기질(ECM)은 세포를 둘러싸고 지지하는 복잡한 네트워크이다. 구성 요소로는 조직의 구조적 강도를 제공하는 콜라겐, 조직의 탄력성을 제공하는 엘라스틴, ECM의 수분 함량과 압축 저항성을 조절하는 프로테오글리칸 등이 있다. ECM은 세포들이 조직 내에서 안정적으로 위치할 수 있도록 기계적 지지를 제공하고, 세포와 상호작용을 하여 세포 분화, 이동, 생존을 조절하는 신호를 전달한다.
이런 과정을 통해 생성된 조직의 주요 유형은 피부와 장 점막 같은 보호, 흡수, 분비 기능을 수행하는 상피 조직, 뼈, 연골, 지방 같은 지지와 결합 기능을 수행하는 결합 조직, 골격근, 심근, 평활근 같은 수축을 통해 운동이 가능한 근육 조직, 두뇌, 척수, 신경 같은 신호 전달 기능을 수행하는 신경 조직 등이다. 이런 조직들이 상호 보완을 하여 상위 조직을 형성하는데 예를 들면 상피 조직과 결합 조직이 어우러져 피부 조직을 형성하는 것이다.
이렇듯 조직 형성은 매우 복잡한 유전자 조절 네트워크가 관여하며, 수많은 유전자와 그 발현을 정교하게 조절하는 다양한 전사 인자, 에피제네틱 메커니즘 등이 포함된다. 또한, 신호 전달, 접착, 이동 등의 세포 간 상호작용이 조화를 이루어야만 조직이 제대로 형성될 수 있다. 이런 정교한 유전자 발현 조절 기능과 세포 간 상호작용이 무작위 돌연변이와 자연선택의 점진적 변화로 형성되었다고 보기는 불가능하다. 왜냐하면, 조직 형성의 중간 형태가 충분한 선택적 이점을 제공하였다면 자연선택에 의해 보존되었겠지만 기능적 중간 형태의 부재는 조직 형성이 점진적 진화의 결과라고 볼 수가 없다. 무엇보다도 조직 형성과 같은 복잡한 구조가 점진적으로 진화했음을 입증하는 중간 단계 화석의 증거가 없다. 그리고, 조직 형성에는 다양한 세포 유형과 조직들이 상호 의존적으로 작동해야 한다. 예를 들어, 근육 조직은 근육세포뿐만 아니라 혈관과 신경 조직도 필요하다. 그런데, 이러한 상호 의존적인 시스템이 동시에 진화하는 것은 매우 낮은 확률이다.
조직이 형성되었으면 조직으로부터 기관이 형성되어야 한다. 이 과정은 서로 다른 유형의 조직들이 상호작용을 하여 복잡한 구조와 기능을 가지는 기관을 만드는 과정이다. 예를 들면, 근육 조직, 결합 조직, 신경 조직 등이 결합하어 심장을 만드는 것이다. 기관이 만들어 지면 기관 내의 세포와 조직이 특정 기능을 수행하도록 더욱 분화되고 전문화되는 기능적 분화가 일어나야 한다. 이렇게 기관이 형성되면 발생학과 세포 생물학의 정교한 조절 메커니즘에 의해 기관이 정해진 자리에 위치하게 된다.
x. 다세포 생명체의 형성
개별 기관이 형성되면, 이들은 단순히 존재하는 것만으로는 충분하지 않다. 모든 기관은 신체 내의 특정 위치에 자리 잡고, 서로 긴밀하게 통합되어야만 하나의 완전하고 잘 기능하는 생명체가 형성될 수 있다. 이러한 통합은 무작위적인 배열이 아니라, 매우 정밀하게 조정된 위치 선정과 연결 과정을 필요로 한다. 각 기관은 신체 내의 다른 기관 및 기관계(예: 순환계, 호흡계, 소화계, 신경계 등)와 기능적으로 적절히 상호작용할 수 있는 위치에 정확히 자리 잡아야 한다. 예를 들어, 심장과 혈관으로 구성된 순환계는 산소를 공급하는 호흡계, 영양분을 전달하는 소화계와 물리적으로 인접해 있어야 하며, 동시에 기능적으로도 적절히 연결되어야 한다. 만약 이러한 연결이 부정확하거나 미흡하다면, 산소와 영양분의 전달이라는 필수적인 기능이 제대로 이루어지지 않아 생명 유지가 불가능해진다. 이 복잡한 배치와 통합 과정에서, 각 기관을 구성하는 조직과 기관 내의 세포들은 끊임없이 각자의 고유한 역할에 맞게 전문화되고 적응해 나간다. 이러한 과정을 기능 분화(functional differentiation)라고 한다. 기능 분화를 통해 생명체의 각 부분은 자신에게 부여된 특정 기능을 보다 효과적으로 수행할 수 있게 되며, 전체 유기체는 하나의 조화로운 시스템으로 작동할 수 있게 된다. 특히 여러 기관과 기관계 간에는 다양한 생화학적 신호와 물질 교환이 이루어지는데, 이 상호작용은 단순한 연결을 넘어서는 고도로 복잡한 체계를 이룬다. 이 복잡한 시스템은 하나라도 빠지거나 미완성된 부분이 존재할 경우, 전체 생명체가 정상적으로 기능할 수 없게 된다. 다시 말해, 생명체는 모든 주요 기관과 기관계가 처음부터 완전한 형태로 존재하고, 서로 정확히 연결되어야만 온전한 기능을 발휘할 수 있다. 이러한 복잡하고 정밀한 상호 의존성은 자연선택, 돌연변이, 또는 점진적 진화 같은 설명만으로는 이해할 수 없는 구조를 보여준다. 왜냐하면 하나의 기관이 부분적으로만 발달했거나, 또는 하나의 기관계가 미완성된 상태로 존재한다면, 생명체는 생존은커녕 기본적인 기능조차 수행할 수 없기 때문이다. 이처럼 생명체를 구성하는 기관들과 기관계는 'all or nothing' 시스템을 이루고 있으며, 이는 처음부터 완성된 상태로 함께 존재해야만 가능한 매우 정교한 설계라 할 수 있다.
지금까지 살펴본 바와 같이, 생명체를 구성하는 주요 요소들이 자연적인 과정만으로 우연히 발생할 수 있을 가능성은, 몇몇 간단한 아미노산을 제외하고는 사실상 전혀 없음을 분명히 알 수 있었다. 오히려 실험적 결과와 이론적 분석은 복잡한 생명체 구성 요소들이 무작위적인 화학 반응으로는 결코 자연적으로 형성될 수 없다는 사실을 더욱 강하게 지지하고 있다. 또한, 아미노산이 결합하여 단백질을 형성하고, 단백질과 핵산이 정교하게 조립되어 DNA와 RNA를 이루며, 나아가 세포, 조직, 기관으로 이어지는 일련의 과정 속에는 단순한 화학 반응을 넘어서는 명확한 방향성과 설계 의도가 존재함이 드러난다. 이 일관된 흐름과 목표성은 무작위적 돌연변이와 자연선택이라는 진화론적 설명으로는 결코 설명될 수 없는 특징이다. 오히려 이는 생명체가 처음부터 고도의 지성과 계획에 의해 의도적으로 설계되었음을 강하게 시사한다. 이제 다음 절에서는, 이러한 관점에서 진화론이 과연 생명체의 기원을 합리적으로 설명할 수 있는지 살펴보도록 하겠다.
b. 진화론은 생명의 기원을 설명할 수 있나?
진화론에서 생명체의 기원을 설명하기 위해 다양한 이론을 제시해 왔는데 그 주요 이론은 자연 선택, 돌연변이, 유전적 부동(genetic drift), 수평적 유전자 이동(horizontal gene transfer) 등이 있다. 각 이론에 대해 간략히 살펴보자.
자연 선택은 유리한 형질을 가진 개체가 더 성공적으로 생존하고 번식하여 여러 세대에 걸쳐 해당 형질이 개체군에서 점점 더 많이 나타나는 과정이다. 자연 선택은 기존에 존재하는 생명체의 변이에 적용되며, 생명체의 기본 구성 요소(아미노산, RNA, 단백질, DNA) 및 구조(세포, 조직, 기관, 다세포 생명체)의 형성에는 적용되지 않기 때문에 생명체의 초기 발생을 설명할 수 없다. 더 나아가, 자연 선택이 작용하려면 이미 재생산과 유전적 변이가 가능한 생명체가 존재해야 하기 때문에 무기물에서 최초의 생명체가 어떻게 형성되었는지를 직접 설명하기는 어렵다.
돌연변이는 유전자의 염기 서열에 무작위적으로 일어나는 변화를 말하며, 자연 선택과 마찬가지로 생명체의 초기 발생과 고도로 조직화된 복잡한 구조를 설명하지 못한다. 보다 더 중요한 문제점은 대부분의 돌연변이가 해롭거나 중립적이다는 것이다. 유익한 돌연변이는 매우 드물기 때문에 이러한 돌연변이가 축적되어 유의미한 변화를 일으키기는 어렵다. 예를 들면, 베시큘러 스토마티티스 바이러스(Vesicular Stomatitis Virus) 연구에서 돌연변이의 결과를 보면 약 40%가 생존에 치명적인 방향으로(fitness=0), 약 31%가 치명적이지 않지만 해로운 방향으로(0
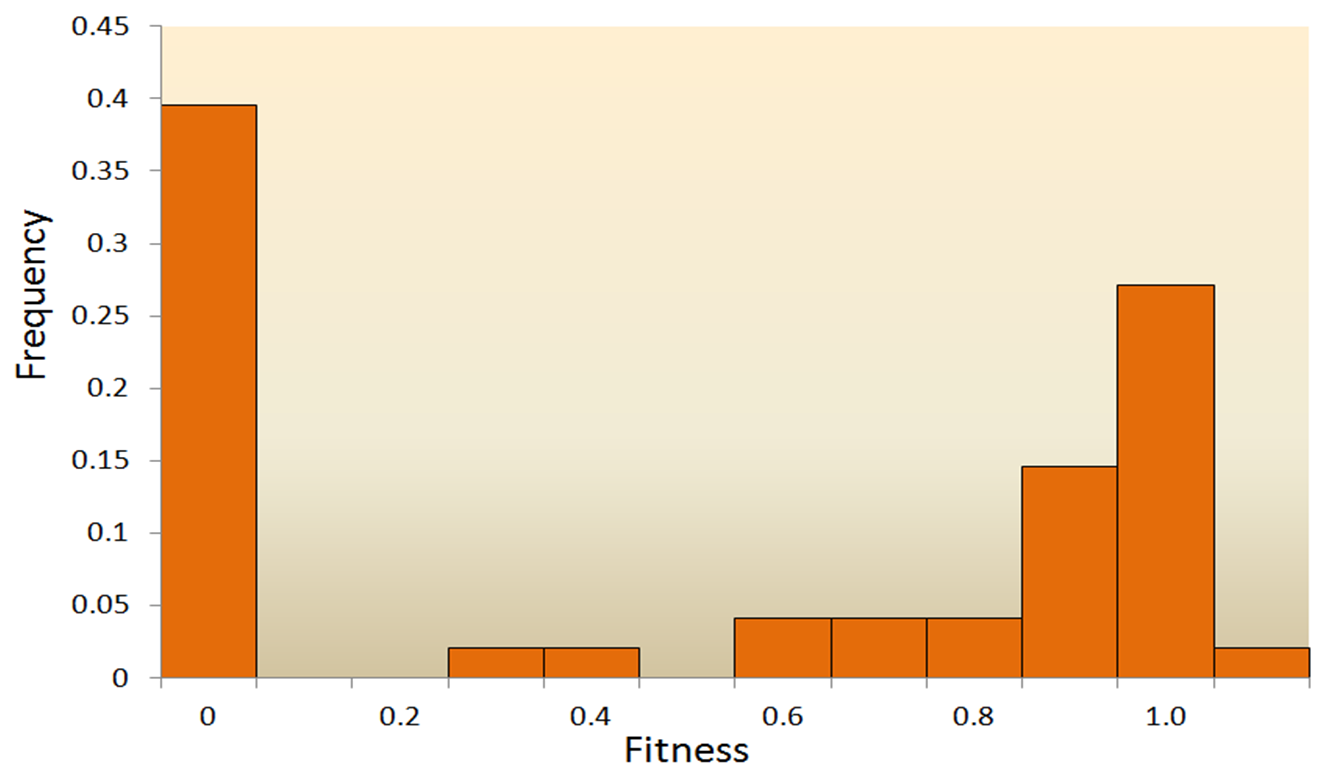
그림 3.6. 돌연변이의 적응도(fitness) 분포
유전적 부동은 유전자 빈도의 무작위 변화를 통해 개체군의 유전적 구성을 변화시킨다는 이론이다. 유전적 부동은 작은 집단에서 더 두드러지며, 대부분의 변화가 발생하는 큰 집단에서는 영향이 적다. 또한, 고도로 조직화한 구조와 시스템의 발달을 설명하기 위한 방향성이 부족하다. 게다가 유전적 부동은 새로운 정보나 기능을 생성하지 못하므로 새로운 특성의 출현이나 복잡한 생물학적 특징의 기원을 설명하지 못한다.
수평적 유전자 이동(HGT)은 주로 원핵생물에 제한된 역할을 하기 때문에 다세포 생물의 복잡한 특성을 설명하지는 못한다. HGT는 외래 유전자를 숙주의 게놈에 통합하기 때문에 유전적 불안정을 초래하여 해로운 돌연변이로 이어질 수 있다. 또한, HGT는 새로운 유전자의 기원을 설명하는 것이 아니라 기존 유전자의 전이를 설명하므로 새로운 특성의 출현을 다루지 못한다.
다음 표는 생명의 기원에 대한 진화 이론의 적용 가능성을 요약한 것이다.
진화론 | 생명의 기원 설명가능? | RNA, 단백질, DNA의 형성 설명가능? | 진화론이 아닌 유전적 적응론인가?* |
자연 선택 | 불가 | 불가 | 예 |
돌연변이 | 불가 | 불가 | 예 |
유전적 부동 | 불가 | 불가 | 예 |
HGT | 불가 | 불가 | N/A |
표 3.2. 생명의 기원에 대한 진화론의 적용 범위(*: 유전적 적응론은 다음 섹션 참조)
위 표에서 볼 수 있듯이, 진화론의 주요 이론들은 지구 생명체의 기원뿐만 아니라 RNA, 단백질, DNA와 같은 생명체의 기본 구성 요소가 어떻게 형성되었는지에 대한 구체적인 메커니즘을 제시하지 못한다. 특히, 생명체의 형성 과정은 고도로 복잡하고 정교한 방향성을 지니고 있음에도 불구하고, 방향성이 없는 무작위적 과정만으로 이를 설명하는 데 한계를 보인다. 그럼에도 불구하고, 대부분의 생물학 교과서 및 《이기적 유전자》와 같은 진화론을 옹호하는 서적들에서는 유전자와 생명체가 원시 지구 환경에서 화학적으로 우연히 생성되었다는 가정을 전제로 삼는다. 하지만 실제로는 철저하게 통제된 실험실 환경에서도 단 몇 가지 아미노산이 합성되는 것이 고작이며, 더욱이 이러한 아미노산이 무작위 과정을 통해 RNA, 단백질, DNA로 형성되는 것은 확률적으로 또는 생화학적으로 전혀 가능성이 없는 일이다. 또한 어떤이는 이렇게 복잡하고 정교한 조건을 요구하는 생명체가 오염되고 혼탁한 심해 열수공에서 탄생되었다고 하는 전혀 받아들이기 어려운 주장을 하기도 한다.
생명체는 변화하는 환경에 대해 끊임없이 적응하며 생존을 유지해 나간다. 이 적응 능력은 이미 생명체의 유전자에 기록이 되어있고 필요에 따라 적절한 유전자 발현되어 환경에 적응한다. 진화론은 단지 변화하는 환경에 생명체가 적응하는 모습을 묘사하는 것에 불과하며 진화론으로 불리기 보다는 "유전적 환경 적응론"으로 불리어야 한다(다음 섹션 참고). 진화론은 이러한 중요한 한계에도 불구하고 과도하게 홍보되어 대부분의 사람은 진화론이 생명체의 기원과 여러 생명체의 형성을 설명하는 이론으로 잘못 알고 있다.
건물을 짓기 위해서는 설계도, 건축 자재, 견고한 기초가 필요하다. 생명체의 형성을 빌딩 건설에 비유하자면 진화론은 설계도(방향성), 건축 자재(RNA, 단백질, DNA), 기초(생명체의 기원) 없이 무작위로 빌딩 건설이 진행되는 것과 같다. 하지만, 빌딩은 설계도 없이 건설될 수 없으며, 잘 설계된 설계도대로 시공되어야만 빌딩이 완공될 수 있다. 빌딩 건설에 설계자가 존재하듯이, 빌딩과는 비교할 수 없이 복잡한 생명체 형성에도 설계자가 반드시 존재해야만 한다.
생명체를 설계하시고 창조하신 분은 바로 창조주 하나님이시다.
c. 다윈의 이론은 진화론인가? 아니면 유전적 적응론인가?
생명체의 진화는 같은 종 사이에서 변이가 일어나는 소진화(Microevolution)와 한 종이 다른 종으로 변하는 대진화(Macroevolution)로 나눌 수 있다. 진화론자들은 자연선택이나 돌연변이에 의한 소진화가 축적되어 대진화가 일어났다고 주장한다. 그러기 위해서는 한 종에서 다른 종으로 옮겨가는 중간종의 존재가 필연적으로 존재하여야 하지만 그 어디를 봐도 중간종은 존재하지 않는다. 다윈 역시 이 사실을 알고 있었고 종과 종 사이의 중간종이 존재하지 않아 자신의 이론을 주장하는 데 어려움이 있다고 고백하고 있다. 종의 기원 6장 ‘진화론의 난관들’ 첫 부분을 보면 이렇게 나온다: “만일 종들이 미세한 점진적 변화에 의해 다른 종들로부터 파생되었다면, 우리는 왜 어디에서나 무수히 많은 전이 형태의 중간종을 보지 못하는가?” 이 사실은 다윈의 딜레마(Darwin’s dilemma)로 알려져 있으며 자신의 진화론이 뭔가 잘못되었다는 것을 시인하는 고백이다.
한때 약 3억 7,500만 년 전에 존재했던 틱타알릭(Tiktaalik)이 물고기와 사족동물의 중간 화석이라는 주장이 있었다. 그러나 Niedzwiedzki 등이 발표한 2010년 Nature 논문을 보면 틱타알릭보다 약 1,800만 년 앞선 사족동물 발자국 화석을 볼 수 있다. 틱타알릭보다 더 오래된 사족동물 발자국의 존재는 틱타알릭이 물고기와 사족동물 사이의 중간종이 아님을 시사한다. 고리 종(ring species)의 개념으로 중간 종의 존재를 설명하려 시도하지만 무리한 주장임을 알 수가 있다. 왜냐하면 고리 종은 종과 종 사이의 중간 종이 아니라 같은 종 내에서 생물학적 교배가 일어나지 않는 종이기 때문이다. 여러 전이 단계의 중간 종에 대한 화석이 없기에 다윈의 이론은 진화론으로 불릴 수 없고 변화하는 생존 환경에 적응하는 생명체의 유전적 적응론으로 불려야 한다. 그 주된 이유는 다음의 밀란코비치 주기와 관련이 있다.
- 밀란코비치 주기
지구는 거의 원 궤도를 돌지만 약 9만 6천 년 주기로 이심률이 변한다. 이심률의 변화는 기후 변화에 영향을 미치며 빙하기와 간빙기의 시기에도 영향을 준다. 그리고 현재 23.5도로 기울어져 있는 자전축의 경사는 41,000년 주기로 21.5도에서 24.5도 사이로 변한다. 이 기울기의 변화는 적도와 극지방 사이의 태양 복사 분포에 영향을 미치며, 계절의 강도를 변화시키고 장기적인 기후 패턴과 빙하기 역학에 중요한 역할을 한다. 지구 자전축은 세차 운동으로 인해 26,000년 주기로 축의 방향이 점진적으로 변한다. 세차운동은 계절의 강도와 시기를 변화시켜 지구 전체 기후 시스템에 영향을 미친다.
이심률, 자전축 기울기, 자전축 세차 운동의 변화가 결합한 효과를 밀란코비치 주기라고 하며, 이 주기는 장기적인 전 세계 기후 변화를 일으킨다. 그 좋은 예가 사하라 사막의 기후 변화다. 태양 복사가 증가하는 기간 동안, 사하라는 더 많은 강우를 경험하여 호수와 강이 있는 무성한 녹색 풍경으로 변하며, 반대로 태양 복사가 감소하면 건조한 조건이 되어 오늘날처럼 광대한 사막이 된다.
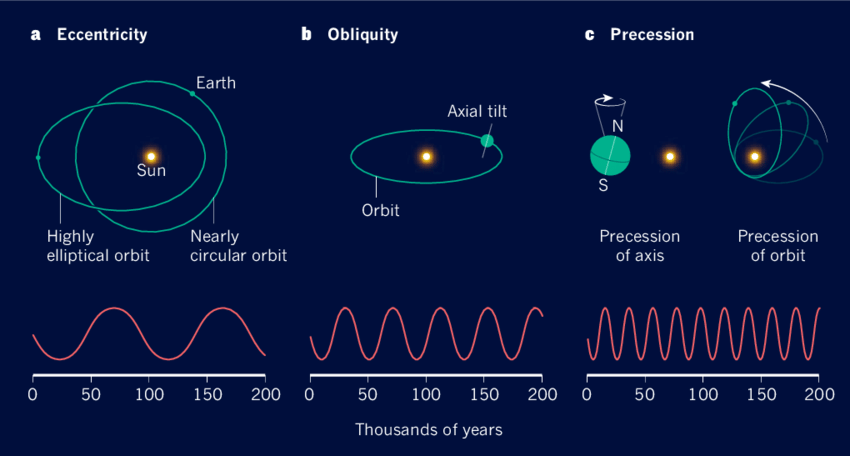
그림 3.7. 밀란코비치 주기의 구성 요소
지구에 이러한 광역적 기후 변화가 발생하게 되면 지구상의 모든 생명체는 유전적 적응을 통해 환경 변화에 몸을 맞춘다. 유전적 적응은 생명체가 멸종되지 않고 오랜 기간 동안 지구에서 생존할 수 있게 해주는 놀라운 생물학적 메커니즘으로, 창조주 하나님께서 생명체의 부호화, 비부호화 DNA 속에 숨겨 놓으신 놀라운 환경 적응력이다. 이런 적응력 때문에 밀란코비치 주기를 겪더라도 생명체들이 소멸되지 않고 지금까지 지구상에서 잘 생존해 왔다. 진화론자들은 이러한 적응력을 진화라는 잘못된 이름으로 부른다. 하지만, 정확하게는 ‘유전자의 환경 적응론’ 또는 ‘유전적 적응론(Theory of genetic adaptation)’으로 불려야 한다. 그렇다면 이제 유전적 적응론의 몇 가지 예를 들어보도록 하자.
- 자외선에 대한 유전적 적응
기후 변화로 인해 인간의 피부가 강한 자외선(UVB, UVA)에 노출되면 여러 단백질과 호르몬이 관여하는 복잡한 메커니즘을 통해 멜라닌 생산이 증가하게 된다. 자외선이 피부 세포의 DNA에 손상을 일으키면 세포의 스트레스와 손상에 대한 반응을 조절하는 중요한 조절자인 p53 단백질이 활성화한다. 활성화된 p53 단백질은 전사 인자로 작용하여 자외선 손상에 대한 보호 반응에 관련된 다양한 유전자의 발현을 촉진한다.
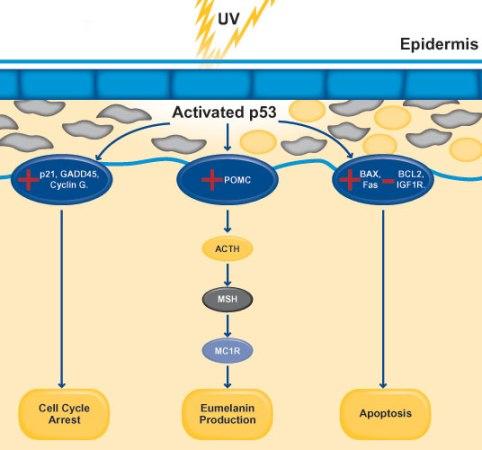
그림 3.8. 유전적 적응에 의한 멜라닌 생성 메커니즘
p53은 POMC라 불리는 호르몬 전구체 단백질 유전자의 발현을 유도하고, POMC는 ACTH라 불리는 부신피질 자극호르몬을 생성하고, ACTH는 MSH라 불리는 멜라닌세포 자극호르몬을 생성하게 된다. 생성된 MSH는 멜라닌세포 표면의 MC1R이라 불리는 멜라노코틴 1 수용체에 결합한다. MSH의 MC1R 결합은 멜라닌세포 내부에서 신호 전달 연쇄 반응을 촉발하고 멜라닌 합성에 관련된 유전자의 발현을 증가시킨다.
멜라닌세포는 멜라닌 생산을 증가시키며, 생산된 멜라닌은 멜라노좀에 포장되어 피부의 외층을 이루는 주요 세포 유형인 각질형성세포로 이동한다. 이동된 멜라닌은 각질형성세포의 핵 위에 보호막을 형성하여 자외선을 흡수하고 분산시켜 피부 세포의 DNA를 추가적인 UV 손상으로부터 보호한다. 이러한 일련의 복잡한 과정들은 생명체가 변화하는 환경에 적응하는 생물학적 메커니즘 중 하나로 유전적 적응이 비교적 짧은 시간 안에 일어날 수 있는 좋은 예이다.
- 북극 환경에 대한 유전적 적응
북극 근처에 사는 이누이트족은 가혹한 북극 환경에서 생존할 수 있도록 유전적 적응을 하였다. 주요 유전적 적응에는 지방산 탈포화효소(FADS) 유전자군의 변이가 포함되어 있는데, 이는 해양 포유류의 전통적인 고지방 식단에서 오메가-3 와 오메가-6 지방산을 대사하는 능력을 향상시킨다. 또한, Carnitine palmitoyltransferase I(CPT1A) 유전자의 유전적 변화는 지방으로부터 에너지 생산을 개선하여 체온 유지에 필수적인 역할을 한다. 이러한 적응은 고지방 식단에도 불구하고 심혈관 질환의 위험을 줄여준다. 더 나아가, 갈색 지방 활동을 조절하는 유전자의 적응은 열 생성을 증가시켜 이누이트족이 극한의 추위에서 열을 생성하고 체온을 유지하는 데 도움을 준다.
이러한 유전적 적응은 최소 20,000년 전, 이누이트 조상들이 러시아와 알래스카 사이의 베링 해협 주변에 살았을 때부터 시작된 것으로 보이는데, 이것은 변화하는 환경에 대한 유전적 적응의 또 다른 좋은 예이다.

그림 3.9. 추운 환경에 유전적 적응한 이누이트족
- 갈색곰에서 북극곰으로의 유전적 적응
갈색곰에서 북극곰으로의 전환은 환경 변화에 의해 유도된 유전적 적응의 다른 예다. 약 40만 년 전, 갈색곰의 한 집단이 북극에서 고립되었고, 이들은 다른 생존 환경에 직면하게 되었다. 시간이 지남에 따라 가혹하고 얼음으로 뒤덮인 환경에서 유리한 유전적 변화가 일어나게 되었는데, 주요 적응에는 지방 대사와 관련된 유전자, 예를 들어 아포지단백질 B(APOB) 유전자의 변화가 포함된다. 이는 그들의 주된 먹이인 물개의 고지방 식단을 처리하는 능력을 향상시켰다. 엔도텔린 수용체 B형(EDNRB)과 Absent in Melanoma 1(AIM1)과 같은 유전자의 적응은 흰색 털의 발달로 이어져 눈과 얼음 속에서 위장할 수 있게 해 주었다. 추가로, 곰의 골격 구조와 사지 형태에 영향을 미치는 유전적 변화는 북극 해역에서 사냥하는 데 중요한 수영 능력을 향상시켰다.
이러한 유전적 적응은 북극곰이 북극 자원을 효율적으로 활용하고 극한의 추위에서 생존하며 갈색곰 조상과 구별되도록 하였다. 중요한 점은 40만 년 동안의 유전적 변화에도 불구하고, 이들은 여전히 곰이며 다른 종으로 변하지 않았음도 주목해야 한다.
그림 3.10. 북극곰의 진화도
- 핀치새 부리의 유전적 적응
다윈의 핀치새 부리 크기와 형태의 변화는 환경 변화에 대한 유전적 적응의 고전적인 예다. 갈라파고스 제도에서 핀치새는 다양한 먹이원을 이용하기 위해 부리 형태를 변화해 왔다. 가뭄이 발생하여 단단한 씨앗이 주요 먹이원이 될 때는, 더 크고 강한 부리를 가진 핀치새가 선택적 이점을 갖게 된다. 반대로, 환경이 부드러운 먹이를 선호하도록 바뀌면, 더 작고 민첩한 부리를 가진 핀치새가 선택적 이점을 갖게 된다. 이러한 적응은 ALX homeobox protein 1(ALX1) 유전자와 같이 부리 형태에 영향을 미치는 특정 유전자와, High-mobility group AT-hook 2(HMGA2) 유전자와 같이 부리 크기에 영향을 미치는 유전자의 변화 결과이다. 환경 변화는 이러한 유전적 변이에 작용하여 다양한 생태적 환경에 적합한 부리 모양을 이끌어 낸다. 여러 세대에 걸쳐 이러한 유전적 적응은 핀치새가 이용 가능한 자원을 효율적으로 활용할 수 있게 하며, 환경적 변화가 어떻게 유전적 변화가 유도하는지 보여 주는 좋은 예다. 핀치새는 약 200만 년 동안 갈라파고스 제도에 살아왔다. 이 긴 기간에도 불구하고, 이들은 여전히 핀치새로 남아 있으며 다른 종으로 변하는 대진화는 일어나지 않았다.
그림 3.11. 변화하는 환경에 유전적 적응을 한 핀치새 부리
다윈은 《종의 기원》에서 그의 이론을 ‘진화론’이라고 명시적으로 주장하지 않았다. 대신, 그는 ‘자연 선택에 의한 종의 기원’을 주제로 ‘변이’, ‘적응’, ‘자연 선택’ 등의 용어를 사용했다. ‘진화’라는 단어는 책의 첫 번째 판이 출간된 후, 동시대 사람들에 의해 널리 사용되자 이후 판에서 명시적으로 사용되었다. 어쨌든 이 ‘진화’라는 용어는 위의 여러 예에서 보여 주었듯이 ‘적응’이라는 단어가 더 적절하기에 다윈의 이론은 ‘진화론’으로 불리기보다는 ‘유전적 적응론’으로 불려야 한다.
d. 우리는 유인원으로부터 진화했을까?
인류학자들에 따르면 인간의 진화는 약 2천 만 년 전 호미노이디아에서 시작되었다고 한다. 호미노이디아는 호미니데와 힐로바티데(긴팔원숭이)로 분화되었고 호미니데는 호미니나이와 퐁기나이(오랑우탄)로, 호미니나이는 호미니니와 고릴리니(고릴라)로, 호미니니는 호미니나와 파니나(침팬지)로, 호미니나는 오스트랄로피테쿠스와 아르디피테쿠스로 분화되었다. 인간은 약 250만 년 전 오스트랄로피테쿠스에서 호모 하빌리스, 호모 에렉투스, 호모 사피엔스를 거쳐 진화했다고 주장한다. 과연 이것이 사실일까?

그림 3.12. 인간은 유인원에서 진화하였나?
앞 장에서 살펴본 대로 핀치새의 경우 200만 년이 흘러도 여전히 핀치새로 남아 있으며 변한 것이라곤 단지 부리 모양뿐이다. 하지만, 거의 같은 기간인 250만 년 동안 침팬지와 같은 유인원이 많은 신체적 변화를 겪고 지능적인 면에서도 비약적인 발전을 이루어 인간으로 진화하였다는 것은 이해하기 어려운 주장이다.
이 문제를 좀 더 자세히 살펴보기 위해 오스트랄로피테쿠스가 인간으로 변하기 위해서는 얼마만큼의 유전자 변이가 필요한지 유전학적으로 접근해 보도록 하자.
그러기 위해서는 오스트랄로피테쿠스와 인간의 유전자 지도가 필요하다. 인간의 유전자 지도는 있지만 오스트랄로피테쿠스의 유전자 지도는 가지고 있지 않다. 오스트랄로피테쿠스 중 가장 잘 알려진 루시의 뇌는 현대 침팬지의 뇌와 비슷한 크기를 가지고 있다. 따라서 오스트랄로피테쿠스의 유전자가 인간과 가장 가까운 침팬지의 유전자와 비슷하다고 가정하자. 침팬지의 유전자 분석은 2005년에 최초로 행하여졌는데 인간의 유전자와 비교해 보면 약 1.2%가 다름이 발견되었다.
부모로부터 다음 세대로 유전자가 전해질 때 약 60개 정도의 유전코드에 에러가 발생하는 돌연변이가 일어난다. 이 숫자는 1억 개의 DNA 문자 중 단지 1개의 문자가 변이되는 확률이다. 한 세대를 25년으로 잡고 오스트랄로피테쿠스로부터 인간까지 진화하는 데 2백 5십만 년이 결린다고 하면 이 기간 동안 10만 세대가 흘렀고 유전자 변이는 10만 곱하기 1억 분의 1인 0.1%의 유전자 변화가 일어날 수가 있다. 이 0.1%(10만/100억)의 유전자 차이는 현생 인류의 여러 인종 간 평균 유전자 차이와 비슷하며 침팬지 수준의 유인원이 인간으로 진화하는 데 필요한 유전자 변이의 약 10%밖에 되지 않는다. 그러므로 2백 5십만 년 동안 오스트랄로피테쿠스부터 인간으로 진화하는 것은 유전학적으로 불가능함을 알 수가 있다. 이 계산은 유전자 돌연 변이가 다음 세대에 이롭게 나타난다는 가정하에 도달한 수치이며 조금이라도 해로운 변이가 함께 나타난다면 인간으로 진화하기에는 더욱더 불가능한 수치에 도달할 것이다.
이 문제는 유전자의 무작위 돌연변이에 의한 코돈 변이를 고려하여 살펴볼 수도 있다. 인간과 침팬지는 모두 약 20,000~25,000개의 단백질 코딩 유전자를 가지고 있다. 각 유전자는 대체 스플라이싱과 번역 후 변형으로 여러 단백질 변이체를 생성할 수 있으며, 그 결과 약 8만~10만 개의 고유한 기능성 단백질을 생성할수 있게된다. 인간 단백질의 아미노산 수는 약 20~33,000개가 된다. 인간과 침팬지 사이에 1%의 유전자 차이가 있다고 하고, 두 종 모두 한 단백질당 평균 100개의 아미노산을 가진 2만 개의 단백질 코딩 유전자를 가지고 있다고 가정하자. 그러면 침팬지가 인간으로 진화하기위해 침판지의 각 단백질당 하나의 아미노산이 인간 아미노산과 일치되게 변이하여야 한다.
침팬지 DNA에서 이러한 돌연변이가 발생하려면 64개의 가능한 코돈 중 어느 코돈으로 변이하여도 되지만 stop 코돈(UAA, UAG, UGA)으로의 변이를 피해야 한다. 왜냐하면 아미노산 배열에 stop코돈이 들어가면 넌센스 돌연변이(nonsense mutation)가 되어 단백질 합성을 멈추고 전혀 기능을 하지 못하는 비기능성 단백질이 되기 때문이다. 2만개의 단백질에서 스톱 코돈과 침팬지 자신의 코돈으로 돌연변이가 일어나지 않고 인간의 코돈에 대응하는 1%의 돌연변이율을 달성할 확률은 (60/64)20000 = 10-561 ≈ 0 이다. 프레임 시프트 돌연변이(뉴클레오타이드의 삽입 또는 삭제)를 고려하지 않더라도 이 확률은 매우 낮으며 무작위적인 우연에 의해 발생하기는 사실상 불가능하다. 이 결과 역시 오스트랄로피테쿠스에서 인간으로의 진화되는것과 같은 대진화는 무작위 돌연변이를 통해서 불가능하다는 것을 알수있다. 따라서 인간은 원숭이로부터 진화하지 않았다.
e. 지적 설계론
지적 설계론(Intelligent Design)은 생명체를 포함한 우주 만물이 우연의 산물이 아니라 지적 설계자이신 창조주 하나님에 의해 설계되었다는 이론으로 창조론의 완곡한 표현이다. 이 때문에 많은 진화론자들이 지적 설계론을 공격하였으며 궁극적으로는 의사 과학으로 매도해 버렸다. 이런 결과로 창조론자들조차 지적 설계론을 받아들이지 않는 사람들이 많다. 하지만 지적 설계는 생명체와 만물의 창조에 대한 과학적 설명으로 창조론의 또 다른 이름으로 받아들여야 한다고 생각한다.
지적 설계와 관련된 주목할 만한 사건은 2005년 미국 펜실베이니아주 도버에서 열린 연방법원 재판이다. 이 재판은 학부모들이 공립학교에서 지적 설계를 가르치는 것이 헌법에 위배된다고 주장하며 소송을 제기하면서 시작되었다. 학부모들은 지적 설계는 본질적으로 종교적 성격을 띠고 있으며 공립학교에서 이를 가르치는 것은 정교분리를 의무화한 미국 헌법의 설립 조항에 위배된다고 주장했다. 재판이 진행되는 동안 지적 설계와 진화론을 지지하는 사람들은 각자의 주장을 펼쳤다. 지적 설계를 대표하는 생화학자 마이클 비히는 생명체의 복잡한 구조는 자연 선택만으로는 설명할 수 없고 지적인 원인에 의해 형성되었음을 제시했다. 그러나 법원은 비히와 다른 지적 설계 지지자들의 주장을 거부하고 대신 진화론 옹호자들의 주장을 받아들였다. 그리고, 판사는 도버 공립학교에서 지적 설계를 가르치는 것이 위헌이라고 판결했다.
이 판결의 가장 큰 문제는 법원이 진화론 지지자들의 주장과 관련 과학 논문을 비판 없이 받아들였다는 데 있다. 이 논문들은 암묵적으로 생명체가 무작위 과정으로 우연히 발생했다고 가정하고 생명체의 환경에 대한 유전적 적응을 진화의 증거로 잘못 해석한 것들이다. 하지만 표 3.2에 요약된 바와 같이 진화론은 현존하는 생명체에만 적용되며 생명의 기원을 설명할 수 없다. 또한 진화론은 생명체의 유전자 코드에 이미 내재된 환경 변화에 대한 유전자의 벌현을 단순히 설명할 뿐이지 그 근원적인 원인은 설명하지 못한다. 그러나 법원은 이러한 사실을 고려하지 않고 판결을 내렸다.
지적 설계론의 시작은 1802년 발간된 윌리엄 페일리의 저서 《자연 신학: 자연의 모습으로부터 수집한 신의 속성 및 존재의 증거》에 나와 있는 시계공 논증으로 거슬러 올라갈 수 있다. 페일리는 시계의 복잡성이 설계자를 암시하는 것처럼 생명과 우주의 복잡성도 신적 창조자를 암시한다고 주장하였다. 지적 설계의 주요 개념에는 특정 복잡성, 환원 불가능한 복잡성, 미세 조정이 포함된다. 미세 조정의 여러 예들은 앞에서 다루었기에 이 장에서는 특정 복잡성과 환원 불가능한 복잡성에 대해 알아보도록 하겠다.
i. 특정 복잡성
지적 설계의 핵심 개념인 특정 복잡성은 자연에서 발견되는 특정 패턴들이 매우 복잡하면서도 특정 기능을 수행하기 위해 정교하게 배열되어 있다는 개념이다. 무작위적인 복잡성과 달리, 특정 복잡성은 단순히 복잡할 뿐만 아니라 특정 결과를 달성하기 위해 질서 있게 배열되어 있다. 이러한 이중적인 특성은 그러한 패턴이 단순한 우연에 의해 발생했기보다는 목적을 가지고 발생했음을 의미한다. 예를 들어 알파벳 1,000개를 무작위로 조합하면 복잡하지만 뜻이 명확하지 않다. 하지만 알파벳 1,000개로 문장을 만들면 복잡한 조합같이 보이지만 그 뜻이 명확하며 어떤 의도를 가지고 그 문장을 작성하였다는 것을 아는 것과 같은 이치다.
특정 복잡성의 대표적인 예는 DNA의 구조이다. DNA의 뉴클레오타이드 서열은 매우 복잡하며, 단일 가닥에서도 수많은 조합을 가지고 있다. DNA 복제 및 복구 메커니즘은 그 복잡성을 더욱 강조한다. 이러한 과정은 유전 정보를 정확하게 복사하고 유지하기 위해 여러 단백질과 효소가 협력하여 작동한다. 뉴클레오타이드 서열은 단순히 복잡할 뿐만 아니라 단백질 합성을 위한 정확한 지침을 암호화할 만큼 매우 구체적이다. DNA 서열의 각 유전자는 특정 단백질에 해당하며, 서열의 작은 변화도 결과 단백질의 기능에 큰 영향을 미칠 수 있다. 또한, DNA는 유전자가 언제 어디서 발현될지를 조절하는 조절 요소를 포함하고 있어 또 다른 특이성을 더한다. 이 사실들은 DNA에서 관찰되는 특정 복잡성이 무작위 돌연변이와 자연 선택과 같은 비방향적인 과정에 의해 발생했을 가능성이 낮다는 것을 의미하며, 대신 지적 설계가 이런 정교하고 기능적으로 특이한 정보의 기원을 더 잘 설명한다.
특정 복잡성의 또 다른 예는 세균 편모다. 세균 편모는 특정 세균이 운동을 위해 사용하는 채찍 같은 모터 구조로, 필라멘트, 갈고리, 기저체와 같은 다양한 구성 요소를 형성하는 약 40개의 다른 단백질로 구성된다. 기저체 자체는 로터, 스테이터, 드라이브 샤프트, 프로펠러를 갖춘 회전 엔진처럼 작동한다. 편모가 작동하려면 이 모든 부품이 존재하고 정확히 조립되어야 하며, 이러한 구성 요소 중 어느 하나라도 없으면 편모는 작동하지 않는다. 편모의 구성 요소는 작동을 위해 매우 특정한 방식으로 배열되어야 하고, 단백질은 정해진 순서대로 조립되어야 하며, 그 형태는 잘 설계된 기계의 부품처럼 정확히 맞아야 한다. 편모는 복잡할 뿐만 아니라 세균을 추진하는 매우 특정한 기능을 수행한다. 놀라운 속도로 작동하며 방향을 바꾸고 에너지 효율적이어서 목적 있는 설계를 나타낸다. 세균 편모의 특정 복잡성은 무작위 돌연변이와 자연 선택에 의해 적절히 설명될 수 없으며, 그러한 고도로 통합되고 기능적인 시스템이 우연히 발생할 가능성은 극히 낮다. 게다가 편모의 중간 형태는 비기능적일 가능성이 높기 때문에 진화론이 제안하는 것처럼 점진적이고 단계적인 진화로는 설명이 불가능하다.
그림 3.13. 세균 편모의 구성
ii. 환원 불가능한 복잡성
환원 불가능한 복잡성은 생화학자 마이클 비히가 도입한 개념으로, 특정 생물학적 시스템이 점진적이고 단계적인 수정으로는 진화할 수 없을 정도로 복잡하다는 이론이다. 이러한 시스템에는 시각 사이클, 세균 편모, 혈액 응고 연쇄 반응, 세포 내 운송 시스템, 항원 항체 반응, 그리고 AMP(아데노신 일인산)의 생합성 등이 있으며, 기본적 기능을 하는 많은 구성 요소가 상호 작용을 하면서 어울려 구성되는 시스템이다. 그리고, 그 구성 요소 중 어느 하나라도 제거되면 그 시스템의 기능이 모두 정지하게 된다. 이러한 정교하고 상호 의존적인 구조는 자연 선택과 무작위 돌연변이만으로는 설명될 수 없으며, 지적 설계자의 설계에 의해서만 가능함을 시사한다.
환원 불가능한 복잡성의 예 중 하나인 시각 사이클(visual cycle)에 대해 자세히 알아보자. 눈의 시각 사이클은 빛을 전기 신호로 변환하여 시각을 가능하게 하는 생화학적 과정이다. 시각 사이클의 주요 구성 요소는 광수용체(간상체와 원추체), 로돕신, 옵신, 레티날, 신호 전달 경로, 신경 처리 과정 등이다. 광수용체는 빛을 감지하는 망막의 세포이며, 간상체는 약한 빛을 감지하고, 원추체는 색을 감지한다. 각 광수용체는 로돕신이라는 광색소 분자를 포함하며, 이는 단백질인 옵신과 빛에 민감한 분자인 레티날로 구성된다. 원추체는 다양한 파장의 빛에 반응하는 다른 옵신을 포함하여 색상 시각을 가능하게 한다. 레티날은 비타민 A의 유도체로, 빛을 흡수하면 모양이 변한다. 이 모양 변화는 옵신을 활성화하여 시각 변환 연쇄 반응을 시작한다. 활성화된 옵신은 트랜스듀신이라는 G-단백질을 활성화하고, 트랜스듀신은 PDE(포스포디에스테라제)를 활성화하여 세포 내 cGMP(고리형 구아노신 일인산) 수치를 낮춘다. cGMP 수치가 낮아지면 광수용체 세포막의 이온 채널이 닫히고, 이는 세포의 과분극을 초래하여 전기 신호를 생성한다. 전기 신호는 양극 세포를 통해 신경절세포로 전달되어 시신경을 통해 뇌로 전송되며, 뇌는 이러한 신호를 처리하여 시각 이미지를 형성한다. 이런 일련의 시각 사이클은 순간적인 속도로 진행되며 아주 적은 빛에 의해서도 효율적으로 진행되어 어두운 곳에서도 물체를 잘 분별할 수 있게 한다. 이해를 돕기위해 이 과정들의 알고리즘을 파이썬 프로그램으로 나타내면 다음과 같다.
컴퓨터 프로그램으로 표현된 시각 사이클
# 초기화: 시각 사이클을 위한 광수용체 환경설정
class VisualCycle:
def __init__(self):
self.photoreceptors = {'rods': [], 'cones': []}
self.initialize_photopigments()
self.signal_pathway_active = False
# 사용자 입력: 들어오는 빛을 감지하고 광색소 활성화 과정 시작
def detect_light(self, light_wavelength):
if light_wavelength in visible_spectrum:
self.activate_photopigment(light_wavelength)
# 이벤트 생성: 레티날의 모양을 변화시키고 옵신을 활성화한 후 신호 전달 경로를 촉발
def activate_photopigment(self, wavelength):
retinal = self.change_retinal_shape(wavelength)
opsin = self.bind_retinal_to_opsin(retinal)
self.start_signal_transduction(opsin)
# 이벤트 처리: 트랜스듀신과 PDE를 활성화하여 cGMP 수치를 감소시키고, 이온 채널을 닫아 전기 신호를 생성
def start_signal_transduction(self, opsin):
self.signal_pathway_active = True
transducin = self.activate_transducin(opsin)
pde = self.activate_pde(transducin)
self.regulate_cGMP_levels(pde)
self.generate_electrical_signal()
# 시그널 처리: cGMP 수치에 따라 이온 채널을 조정하여 전기 신호 생성을 촉진
def regulate_cGMP_levels(self, pde):
cGMP_level = self.reduce_cGMP(pde)
self.adjust_ion_channels(cGMP_level)
# 시그널 출력: 전기 신호를 생성하고 뇌로 전송
def generate_electrical_signal(self):
if self.signal_pathway_active:
electrical_signal = self.create_signal()
self.transmit_signal_to_brain(electrical_signal)
# 네트워크 통신: 신호를 처리하고 양극 및 신경절세포를 통해 전달, 궁극적으로 시신경을 통해 신호 전송
def transmit_signal_to_brain(self, signal):
bipolar_cells = self.process_signal_with_bipolar_cells(signal)
ganglion_cells = self.forward_signal_to_ganglion(bipolar_cells)
optic_nerve = self.send_signal_via_optic_nerve(ganglion_cells)
self.visual_perception(optic_nerve)
# 최종 출력: 뇌가 신호를 해독하고 처리하여 시각 이미지를 생성
def visual_perception(self, optic_nerve):
visual_cortex = self.decode_signal(optic_nerve)
self.render_image(visual_cortex)
시각 사이클을 컴퓨터 프로그램으로 나타낸 위 비유에서 우리는 다음과 같은 사실을 명확히 알 수가 있다. 시각 사이클의 각 구성 요소는 상호 의존적이며, 광수용체, 로돕신, 레티날, 트랜스듀신, PDE, 이온 채널이 이미 존재하고 제대로 기능해야 한다. 만일 어느 한 구성 요소라도 존재하지 않거나, 시각 사이클의 순서가 바뀌어도 시각 사이클은 작동하지 않는다. 따라서 이러한 복잡한 시스템은 모든 구성 요소가 없는 중간 단계는 기능하지 않으므로 작은 점진적 변화의 연속을 통한 진화론으로는 설명할 수 없다. 시각 사이클이 컴퓨터 프로그램으로 잘 표현되는 것은 눈이 창조주에 의해 지적으로 설계되었다는 것을 의미한다. 이것이 지적 설계론의 한 예이며 눈의 설계도는 창조주께서 설계하신 염색체 11번에 있는 PAX6라는 눈을 만드는 유전자이다.
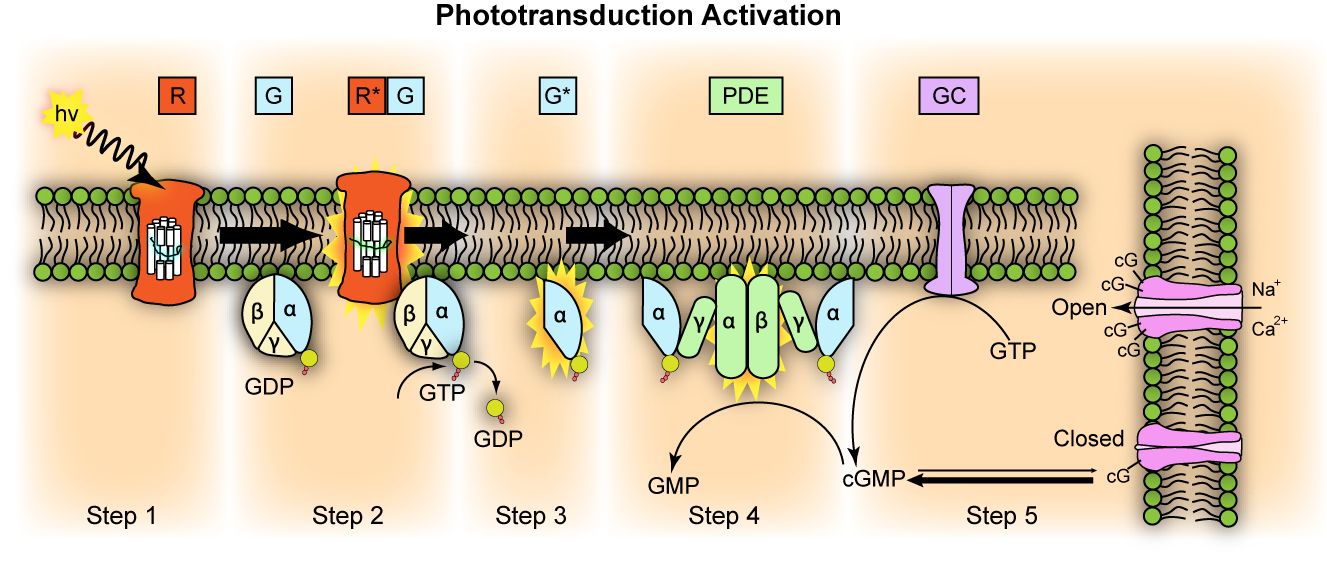
그림 3.14. 시각적 사이클의 작동 과정
그럼에도 불구하고 진화론자들은 눈의 기원을 설명하기 위해 충분한 근거가 뒷받침되지 않은 가설을 제시한다. 다윈은 무척추동물에서 발견되는 단순한 시신경이 점진적인 변화를 거쳐 척추동물의 복잡한 눈으로 진화했다고 주장했다. 도킨스 역시 스웨덴 생물학자들의 모의실험을 언급하면서 눈은 명암과 방향 정도를 감지하는 단순한 조직에서 36만 4000세대를 거쳐 수정체를 갖춘 카메라 수준으로 진화하였다고 주장한다. 위의 실험은 어류의 눈을 대상으로 한 것인데 만일 어류의 한 세대를 10년으로 잡는다면 시신경에서 완전한 눈으로 진화하기 위해 약 360만 년이 걸린다는 이야기다. 이 긴 기간 동안 보이지 않는 눈으로 어떻게 먹이 활동을 하였으며 포식자들로부터 어떻게 살아남았는지 의문이다. 그리고, DNA나 단백질조차 자연적으로 형성될 수 없는데 최초의 시신경은 어떤 과정을 거쳐 생성되었는지에 대한 설명이 없다. 또한, 눈의 진화가 가능해지려면 시각에 관련된 수많은 유전자 변이와 단백질의 상호작용이 있어야 한다. 그런데 단순한 시신경에서 수정체를 갖춘 눈으로 발전하기까지 이러한 유전자 네트워크가 어떤 과정을 거쳐 점진적으로 형성되었는지에 대한 분자생물학적 증거가 부족하다. 예를 들어, 로돕신, 레티날, 트랜스듀신, PDE, 이온 채널, 신경 회로망 등의 요소가 개별적으로 진화했다고 하더라도 이들이 조화를 이루며 기능을 발휘하려면 동시에 특정 방식으로 상호작용을 해야 한다. 그런데, 이러한 복잡한 요소들이 무작위적 과정을 통해 적절하게 취합되어 정교한 시각 기능을 형성하였다는 주장은 논리적으로 납득하기 어렵다.
iii. 지적 설계론 주요 도서
이 장에서는 지적 설계론에 대한 중요한 서적들을 소개하고자 한다.
위기에 처한 진화론(Evolution: A Theory in Crisis, Michael Denton, 1985): 이 책에서 마이클 덴튼은 분자 생물학과 고생물학 등 여러 분야의 증거를 제시하며, 생명체에서 관찰되는 정교한 구조와 기능이 무작위 돌연변이와 자연 선택으로는 설명하기 힘들며 지적 설계를 가리킨다고 설명하고 있다.
다윈의 블랙박스(Darwin's Black Box: The Biochemical Challenge to Evolution, Michael J. Behe, 2006): 이 책에서 마이클 비히는 환원 불가능한 복잡성 개념을 소개하며, 세균 편모와 같은 특정 생물학적 시스템이 자연 선택이나 점진적 진화로는 설명할 수 없을 정도로 복잡하며 지적 설계의 산물이어야만 설명 가능하다고 주장한다. 이 책이 미국에서 처음 나왔을 때 Nature나 Science와 같은 과학적인 저널은 물론이고 Wall Street Journal과 같은 비교적 대중적인 저널에 이르기까지 많은 곳에서 비평되었다. 비히의 책은 창조론을 지적 설계라는 과학의 경지로 끌어 올렸다는 평가를 받는다.
심판대의 다윈(Darwin on Trial, Phillip Johnson, 2010): 필립 존슨에 의해 쓰인 심판대 위의 다윈은 지적 설계 운동의 출발점이며 다윈주의의 학술적 비평과 새로운 과학 패러다임으로서 지적 설계 운동을 열었다는 평가를 받고 있다. 이 책의 제1장은 법적 배경으로서 이 책의 저술 목적을 피력하고 있고, 제2장부터 제8장에 이르기까지 자연선택, 크고 작은 돌연변이, 화석 문제, 진화의 사실, 척추동물 계통, 분자적 증거, 생명 이전의 진화 등 진화론과 관련된 직접적 내용을 다루며, 제9장부터 제12장까지는 과학의 규칙, 다윈주의 종교, 다윈주의 교육, 과학과 의사 과학 등 진화론과 관련된 철학적 내용을 다루고 있다.
세포 속의 시그니쳐: DNA와 지적 설계의 증거(Signature in the Cell: DNA and the Evidence for Intelligent Design, Stephen C. Meyer, 2010): 이 책은 세포에 대한 현대 분자생물학과 생화학의 발견에 기초하여 지구상 최초 생명의 출현은 지적 존재의 설계에 의해서 가능했다는 것을 논증하고 있다. 그는 DNA 내 유전 정보의 정교함을 바탕으로 지적 설계에 대한 상세한 사례를 제시하며, 생명의 기원이 무작위 과정보다는 목적 있는 창조를 가리킨다고 제안한다.
다윈 퇴보: 진화에 도전하는 DNA에 대한 새로운 과학(Darwin Devolves: The New Science About DNA That Challenges Evolution, Michael J. Behe, 2020): 비히의 또 다른 책으로 최근의 유전적 발견들이 전통적인 다윈의 진화론을 약화한다고 주장한다. 그는 자연 선택과 무작위 돌연변이가 소규모 적응을 설명할 수는 있지만, 세포 내의 분자 기계를 설명하기에는 부족하다고 주장한다. 비히는 “퇴보”라는 개념을 도입하여 돌연변이가 새로운 유익한 특성을 창출하기보다는 유전 정보를 잃게 만들며, 이러한 유전적 한계가 지적 설계자의 필요성을 가리킨다고 주장하고, 지적 설계가 생명의 복잡성을 설명하는 더 적합하다고 제안한다.
생명 기원의 미스터리: 현재 이론들의 재평가(The Mystery of Life’s Origin: Reassessing Current Theories, Charles B. Thaxton et al., 2020): 이 책은 생명의 기원을 설명하는 다양한 진화론적 이론의 단점을 논의하며 지적 설계를 과학적으로 유효한 대안으로 소개하고 있다. 현대 지적 설계 운동의 기초를 마련한 책으로 평가된다.
설계 추론: 작은 확률을 통한 우연의 배제(The Design Inference: Eliminating Chance through Small Probabilities, William A. Dembski & Winston Ewert, 2023): 이 책은 자연계에서 지적 설계를 탐지하기 위한 이론적 기초를 제공한다. 특정 복잡성, 작은 확률, 필터링 기법의 3가지 주요 개념을 사용하여 지적 설계를 추론해 낸다. 어떤 현상이 복잡하면서 동시에 특정한 기능을 수행할 때, 또는 자연 현상이 무작위적으로 발생할 확률이 매우 낮을 때는 지적 원인에 의해 설계되었음을 나타낸다. 필터링 기법은 특정 현상이 자연법칙에 의해 설명될 수 있는지를 확인하여, 만약 자연법칙으로 설명될 수 없다면, 두 번째 단계에서는 무작위적인 과정에 의해 설명될 수 있는지를 검토한다. 무작위적인 과정으로도 설명되지 않는다면, 마지막 단계에서 지적 설계에 의해 설명될 가능성을 고려하는 것이 필터링 기법이다. 복잡한 생물학적 시스템은 무작위적인 과정에 의해 형성될 가능성이 희박하기 때문에 지적 원인에 의해 설계되었음을 나타낸다.
f. 입자 물리학과 창조론
앞에서 우리는 원자, 아미노산, RNA, 단백질, DNA, 세포 등으로 구성된 생명체의 기원에 대해 살펴보았다. 그리고 이 논의에서 생명체의 구성하고 있는 최소 단위인 원자는 당연히 존재하고 있음을 가정하고 출발하였다. 그렇다면, 천지 만물을 구성하고 있는 원자는 어떻게 존재하게 되었을까? 단순한 우연의 결과일까, 아니면 창조주에 의해 창조된 것일까? 여기서는 천지 만물의 근원인 원자들을 구성하고 있는 아원자(소립자)들은 어떻게 생성되었는지 알아보도록 하겠다.
입자 물리학의 표준 모형에 의하면 이 세상의 모든 물질은 6개의 쿼크, 6개의 렙톤, 4개의 게이지 보손(글루온, 광자, Z 보손, W 보손), 힉스 보손 등 17개의 기본 입자로 이루어져 있다. 각 입자는 특정한 질량, 전하, 스핀을 가지고 있으며 마치 세포 속에서 각자 맡은 역할을 성실히 수행하는 세포소기관들처럼 입자들의 상호작용에 있어 특정한 역할을 수행한다.
쿼크는 물질을 구성하는 중요한 뼈대로 2개의 업쿼크와 1개의 다운쿼크가 강력으로 묶여 있으면 양성자가 되고, 1개의 업쿼크와 2개의 다운쿼크가 강력으로 묶여 있으면 중성자가 된다. 강력은 쿼크들이 글루온을 주고받으면서 생성되며, 거리의 역 제곱에 비례하는 중력이나 전자기력과는 달리 핵자 내에서는 거리가 멀어지면 힘이 세지고 거리가 가까워지면 힘이 약해져 쿼크들이 일정한 거리를 유지하게 만든다. 쿼크들은 입자들의 상호작용 중 다른 쿼크들로 바뀌는데, 예를 들면 중성자가 베타 붕괴로 양성자로 바뀌면 중성자에 있던 다운쿼크가 업쿼크로 바뀌면서 양성자가 된다. 이렇게 쿼크가 바뀌는 패턴은 수학적으로 CKM 행렬에 따른다.
기본 입자들은 힉스 메커니즘에 의해 질량을 부여받는데 아무 질량 값이나 받는 게 아니라 각 입자마다 꼭 정해진 질량 값을 부여받는다. 입자들이 질량을 부여받을 때 힉스 입자가 생성되는데 이 입자의 실체가 2012년에 확인되었고 이 발견으로 힉스와 앙글레르가 노벨상을 받았다.
게이지 보손은 입자들의 상호작용 중 힘을 매개하는데 광자는 전자기력, W 보손과 Z 보손은 약력, 글루온은 강력을 매개한다. 이 입자 간의 상호작용은 양자역학, 양자장론, 양자 색역학 등을 통해 모두 수학적으로 기술될 수 있다.
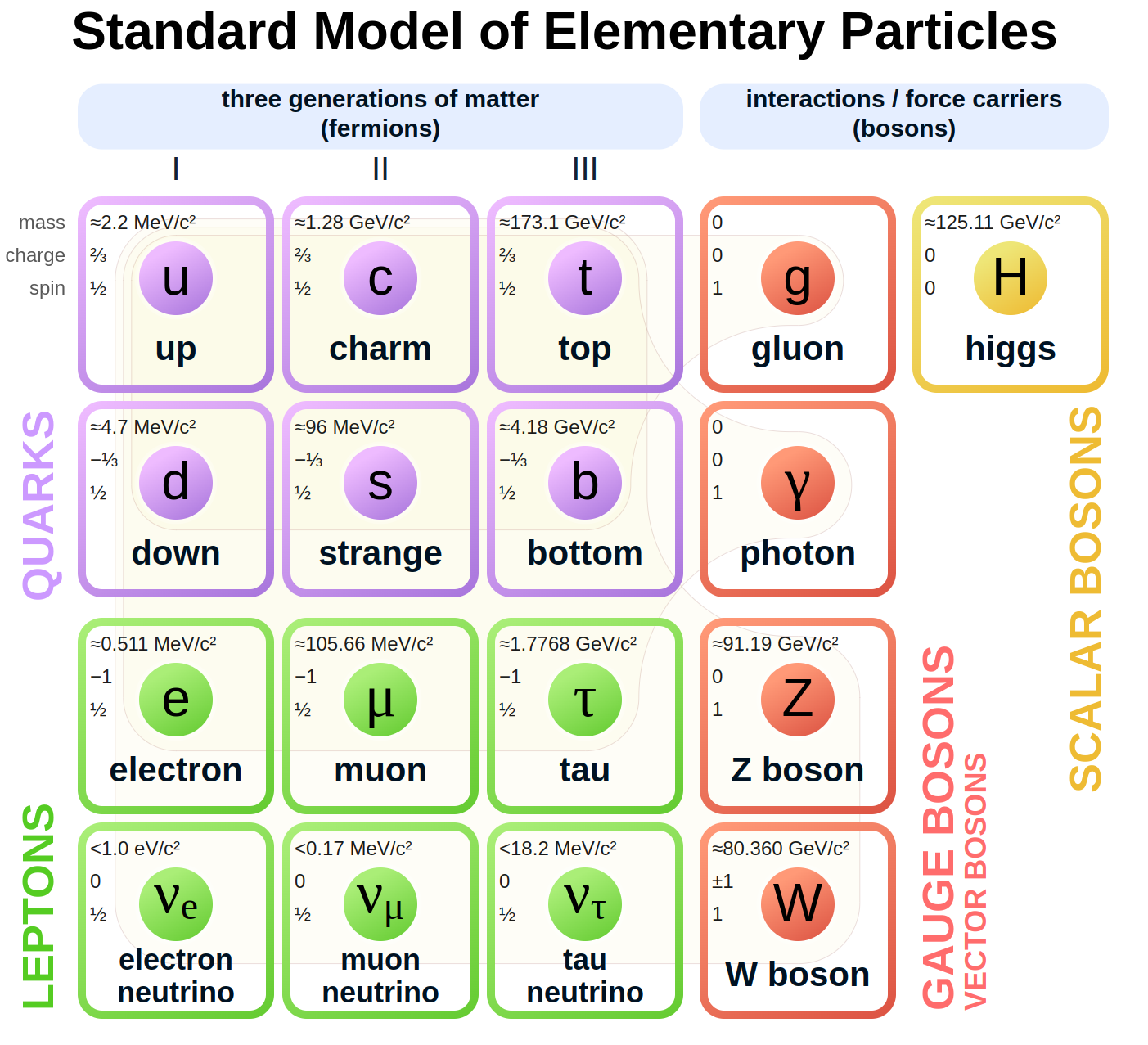
그림 3.15. 표준 모형의 기본 입자들
입자 물리학의 세계는 광대하고 조금 난해하지만 이상의 간단한 사실들만 놓고 볼 때 다음과 같은 근원적인 질문을 던질 수 있다.
- 만물을 구성하는 17개의 기본 입자는 어떻게 만들어졌을까? 에너지에서 우연히 추출되었을까? 아니면 정밀한 수학적 설계에 따라 생성되었을까?
- 게이지 보손들은 어떻게 하여 힘을 매개하는 특성을 획득하였는가? 우연히 이런 메커니즘이 생성된 것인가? 아니면 정밀한 수학적 설계의 산물인가?
- 기본 입자마다 정해진 질량을 부여하는 힉스 메커니즘은 자연적으로 생겨났는가? 아니면 정밀한 수학적 설계의 산물인가?
- 베타 붕괴로 중성자가 양성자로 바뀌는 과정은 자연적인가? 아니면 수학적 설계의 산물인가?
- 어떻게 해서 입자들의 모든 상호작용이 수학적으로 기술 가능한 것인가? 우연인가 아니면 수학적 원리에 따라 설계되었기 때문인가?
이 5개의 사실 중 어느 하나라도 무작위 프로세스에 의한 우연의 결과라면 이 세상은 존재하지 않았을지 모른다. 왜냐하면 기본 입자 중 하나라도 빠져 있거나, 힉스 메커니즘이 확립되지 않았거나, 기본 입자의 질량과 스핀 값이 조금이라도 달랐다면 전자, 양성자, 중성자 등이 만들어지지 않았을 것이고, 혹시 만들어졌더라도 서로 단단하게 결합하지 못하였을 것이다. 그랬다면 세상의 모든 물질은 붕괴하어 인간을 포함한 그 어떤 것도 형성될 수 없었을 것이다. 여기서 우리는 입자 물리학의 세계에서도 지적 설계론의 환원 불가능한 복잡성을 관찰할 수가 있다.
물질을 형성하기 위한 기본 입자의 생성은 생명체 형성을 위한 세포와 세포소기관의 생성에 비유할 수 있다. 특정 세포와 세포소기관이 각각 고유한 역할과 특성을 가지고 있어 생명체를 만들고 유지하는 것처럼, 기본 입자들도 각각 고유한 역할과 특성을 가지고 있어 원자, 분자, 그리고 궁극적으로는 물질을 형성하고 유지한다. 이런 유사점은 세포의 미시적 수준, 물질의 소립자 수준, 생명체, 별, 은하 등의 거시적 수준 등 모든 자연계에 지적 설계의 개념이 들어 있는 것을 알 수가 있다.
기본 입자들의 존재와 상호 작용들이 복잡한 양자역학의 수식으로 표현될 수 있다는 것은 이 모든 것들이 수학적으로 정교하게 설계되었다는 것을 뜻한다. 그렇지 않을 경우 각각의 기본 입자들이 생각하는 능력이 있어 어떻게 하면 물질을 구성할 수 있을지 알아서 정확한 질량, 전하, 스핀 값들을 가지고 생성되며, 서로의 상호작용에서 복잡한 수학적 원리를 사용하여 행동한다는 것을 받아들여야 한다. 이 사실은 사고 능력이 없는 기본 입자들이 어려운 양자 역학의 원리를 완벽히 이해하고 적용하고 있다는 것을 받아들여야 하는데, 이것이 사실이 아니라는 것을 누구라도 알 것이기 때문이다.
g. 외계인과 창조론
우주에 있는 수많은 별 중에는 우리 태양계와 비슷한 조건을 가진 별들이 셀 수 없이 많으며, 그런 별 주위를 도는 행성중 우리 지구와 비슷한 조건을 가진 행성들 역시 무수히 많을 것이다. 따라서 진화론이 맞다면 우리 지구에만 생명체가 발생한 것이 아니라 지구와 비슷한 조건을 가진 수많은 행성에서도 생명체들이 발생하어야 할 것이다. 그럴 경우 어떤 행성에서는 이제 막 생명체가 발생하여 박테리아 수준의 생명체가 살고 있을 것이고 어떤 행성에서는 오래전에 생명체가 발생하여 현재 우리가 상상할 수도 없는 고도의 문명을 누리고 있을 것이다. 만일 우리가 이렇게 많은 우주 생명체의 존재를 발견한다면 창조론과 진화론의 논쟁에 마침표를 찍을 수 있을 것이다.
은하계 내 외계 생명체가 살 수 있는 행성의 수 N은 다음 드레이크 방정식을 통해 추정할 수 있다:
N = R* × fp× ne× fl× fi× fc× L
여기서, R*은 별 탄생률, fp는 별이 행성을 가진 확률, ne는 생명체가 나타날 수 있는 행성의 수, fl 은 실제로 생명체가 탄생할 확률, fi는 생명체가 지적 생명체로 진화할 확률, fc 는 우주로 신호를 보낼 수 있는 문명의 비율, L은 그 문명이 지속할 기간이다. 각 변수에 적절한 값을 적용하면 한 은하당 외계 문명이 존재할 행성은 약 2개로 추정된다.

그림 3.16. 외계인이 존재하는가?
우주에 외계인의 존재 여부를 직접적으로 알아보는 외계인 탐사 계획(SETI 계획)은 1960년대 초 드레이크에 의해 시작되었다. 초기에 행해진 외계인 탐사 계획은 오즈마 계획과 오즈마 계획 II로 불리는데 미국 그린뱅크에 있는 전파망원경을 사용하여 지구에서 가까운 약 650여 개의 태양형 별들을 관측하였다. 그 후 나사에서 수행된 사이클롭스 계획, 그 후속으로 수행된 피닉스 계획, 버클리 대학에서 수행된 세렌딥, SETI@home, 알렌 전파망원경 계획, 하버드와 프린스튼 대학에서 수행된 광학 세티 계획 등을 통해 태양과 비슷한 1천여 개의 별들이 더 관측되었다. 그럼 그 결과는 어떻게 되었을까?

그림 3.17. SETI에 사용된 전파 망원경
지금까지 60여 년 동안 여러 세티 계획을 통해 외계인의 존재를 추적하였지만 그 존재를 발견하지 못했다. 최근 백만 개의 별과 백 개의 은하를 관측하고 인공지능으로 분석한 ‘획기적 청음(Breakthrough Listen)’ 계획의 결과도 역시 외계문명을 발견하지 못하였다. 그렇다면 외계인은 존재하지 않는 것인가?
외계인의 존재는 확인되지 않았지만 만일 수많은 외계 문명이 존재한다면 그들은 이미 우리를 방문했거나 지금 방문하고 있을지도 모른다. 그럴 경우 그들은 어떤 방법으로 우주여행을 할까? 우리가 일반적으로 생각하는 UFO와 같은 비행 물체를 타고 광대한 우주를 여행한다는 것은 거의 불가능하다. 왜냐하면 비행기보다 30배나 빠른 로켓으로 비행해도 가장 가까운 별인 프록시마 센타우리까지 가는데 15만 년이 걸리기 때문이다. 이렇게 광활한 우주를 단시간에 여행하는 방법을 찾아야 하는데 가능한 방법으로는 워프 드라이브나 웜홀을 통한 여행이다.
워프 드라이브는 아인슈타인의 일반 상대성 이론에서 영감을 받아 빛보다 빠른 우주 여행을 위한 이론적 개념이다. 1994년 물리학자 미구엘 알쿠비에르가 제안한 워프 드라이브는 우주선 앞의 공간을 수축시키고 뒤의 공간을 확장하는 '워프 버블'을 생성하여 상대론을 위반하지 않고도 외부 관측자에 비해 빛보다 빠르게 움직일 수 있는 방법이다. 이론적으로는 가능한 방법이지만 실재적으로 사용하기 위해서는 음의 에너지 밀도를 가진 물질이 필요하기에 현재로서는 실현 가능성이 희박하다.
그림 3.18. 웜홀
아인슈타인의 일반 상대성 이론에 의해 예측된 웜홀 또는 아인슈타인-로젠 다리는 광대한 우주 거리를 웜흘을 통해 순간적으로 여행할 수 있게 해줄 수 있다. 하지만 실제로 이 방법을 사용하려면 통과하는 웜홀을 안정화해야 하며, 웜흘의 붕괴를 막기 위해 워프 드라이버처럼 음의 에너지 밀도를 가진 물질이 필요하다.
우주를 순간적으로 이동하기 위한 또 다른 방법은 teleportation으로 불리는 순간이동 또는 공간이동이다. 막 우주론(Brane cosmology)에 의하면 우리의 3차원 우주는 보다 높은 차원인 초공간(bulk 또는 hyperspace)의 일부라는 것이다. 초공간을 통한 순간이동은 기존의 3차원 공간을 우회하여 광대한 거리를 순간적으로 이동할 수 있는 방법이다.

그림 3.19. 공간이동
최근의 관측에 의하면 최초의 은하와 별들은 지금으로부터 약 134억 년 전에 형성되었는데 진화론이 맞을 경우 이미 그때부터 생명체들의 진화가 시작되었을 것이다. 따라서 어떤 생명체는 우리보다도 1억 년, 10억 년, 또는 최고 94억 년(우리는 40억 년 전부터 진화를 시작하였음) 먼저 진화를 시작하여 지금은 우리가 상상하지 못할 정도의 고도 문명을 이루고 있을 것이다.
이런 생명체들이 134억 년 전부터 우주의 여러 곳에서 균일한 빈도로 진화를 시작하였다고 가정하면 약 70%(94억 년/134억 년) 정도의 우주 생명체들이 우리보다 먼저 진화를 시작하였고 그중 90%는 우리보다 9억4천만 년 먼저 진화하였고 99%는 우리보다 9천4백만 년 더 일찍 진화를 시작하였을 것이다. 이제 극히 보수적인 입장을 취하여 그들 중 99%가 아니라 1%만, 또한 우리보다 9천4백만 년이 아니라 단지 100만 년 먼저 진화하였다고 가정해 보자.
그럴 경우 앞의 드레이크 방정식에서 계산된 4천억 개의 지적 생명체 중 1%인 40억 개의 지적 생명체가 우리보다도 백만 년 먼저 진화를 시작하였을 것이다. 그들이 우리보다 100만 년 전 우리와 비슷한 진화 경로를 따라 진화했다고 가정하면 그들의 문명은 우리 문명보다 100만 년 더 발전했을 것이다. 이렇게 100만 년 먼저 진화한 외계 문명체는 우주여행을 위한 순간이동 방법을 이미 발견하였을 것이고 지금쯤은 우리가 이웃을 방문하는 것처럼 우주 어디든 순간적으로 여행할 수 있을 것이다.
그러한 문명체의 한 문명당 10억 명의 외계인이 살고 있다고 하면 지구를 이웃집처럼 순식간에 방문할 수 있는 전체 외계인 수는 40억 개의 지적 생명체 × 10억 명으로 400경 명이 됨을 알 수 있다. 그들 중 모두가 한 번씩 지구를 방문한다면 400경 명, 1%만 방문한다면 4,000조 명의 외계인들이 우리 지구를 방문할 수 있을 것이다. 만일 그들이 하루에 한 번씩 지구를 방문한다면 4,000조 명의 외계인이, 일 년에 한 번씩이면 11조 명, 십 년에 한 번씩이면 1조 천억 명의 외계인들이 매일 지구를 방문하게 될 것이다. 그렇게 될 경우 우리 지구는 지구에 사는 인간들보다 훨씬 더 많은 외계인들이 몰려와 그야말로 발 디딜 틈이 없을 정도의 장사진을 이룰 것이다. 그런데 과거에도 그리고 현재에도 그런 외계인의 방문 흔적이 전혀 없는 것은 어떻게 이해하여야 할까?
외계인의 지구 방문 흔적이 없는 것에 대해 1950년에 엔리코 페르미가 의문을 가졌고 “모두 어디에 있지?”라고 물었던 페르미 역설(Fermi paradox)로 알려져 있다. 이에 대한 설명으로서는 i) 외계인은 벌써 우리 곁에 와 있지만 우리가 알지 못할 뿐이다, ii) 외계인은 존재하지 않고 우주에는 우리뿐이다. i)의 경우 세계 여러 관광지에서 지구인들보다 훨씬 더 많은 외계인을 볼 수 있을 것이고, ii)의 경우 외계인의 수를 추정한 드레이크 방정식에서 진화론에 근거한 ‘행성에서 생명체가 자연적으로 생겨날 확률’이 제로가 될 것이다. 외계인이 박테리아나 투명 인간이 아니라면 어떤 형태로든 우리에게 그들의 존재가 드러났을 터인데 아직까지 그들의 존재를 확인하지 못했다는 사실은 드레이크 방정식에서 가정한 진화론이 맞지 않음을 시사한다.
h. 생명체의 본능과 창조론
컴퓨터는 하드웨어와 소프트웨어로만 구성되어 있는것이 아니라 그 중간 형태인 펌웨어(Firmware)도 포함한다. 펌웨어는 컴퓨터를 켤 때 시작되는 BIOS 또는 UEFI 프로그램으로 컴퓨터의 하드웨어 및 입출력 장치들을 통제하는 프로그램이 들어 있다. 즉 컴퓨터를 만든 사람이 그 컴퓨터가 어떻게 작동되어야 할지 지시하는 사항이 하나하나 기록되어 있는 것으로, 생명체가 자연에서 어떻게 생존해야 하는지 기록되어 있는 생명체의 본능과 같은 역할을 수행한다고 볼 수 있다. 컴퓨터 설계자가 ROM에 펌웨어를 내장한 것처럼, 본능은 창조주께서 생명체의 두뇌와 신경계에 내장한 것으로 생명체가 무작위의 결과로 만들어졌기 보다는 컴퓨터와 같이 창조주에 의해 창조되었음을 시사한다. 그러면 생명체에 내재된 이 본능이 생존을 위해 어떻게 이행되는지 큰미장이 꽃벌과 위버새(Weaver bird)의 집 짓기를 통해 살펴보기로 하자.
i. 큰미장이 꽃벌의 집 짓기
큰미장이 꽃벌의 집 짓기는 파브르 곤충기에 자세히 나와 있는데 내용을 정리하면 다음과 같다. 큰미장이 꽃벌은 강가 모래밭의 평평한 돌을 고른 후 그 위에 침으로 반죽한 흙덩이를 날라와 도넛 형태로 쌓으며 튼튼한 집을 짓는다. 그 후 지어진 집에 꿀과 꽃가루를 채우고 알을 하나 낳은 후 집 입구를 막아 버린다. 여기서부터 파브르는 재미있는 실험들을 진행한다.
벌이 집 짓기의 첫 단계인 흙도넛을 쌓아 기초를 만든 후 다시 흙덩이를 가지러 갔을 때 집 짓기를 거의 마친 다른 벌의 집과 맞바꾸어 놓으면 흙덩이를 물고 돌아온 벌이 그것을 보고 어떻게 행동할까? 그럴 경우 흙덩이를 버리고 거의 완성된 집에 꿀을 채우는 것이 정상적이 행동이겠지만 그렇게 하지 않고 완성된 집 위에 가져온 반죽을 발라 집 짓기를 계속한다.
이와는 반대로 집이 완전히 지어져 꿀이 저장되고 있는 집을 벌이 없을 때 기초공사만 마친 집과 맞바꾸어 놓았다면 꿀을 모아 집으로 돌아온 벌은 어떻게 행동할까? 가져온 꿀을 내려놓고 대신 흙반죽을 가져다가 집을 완성시키기보다는 금방 넘칠 정도의 낮은 집에다가 계속해서 꿀을 저장한다.
그림 3.20. 완성된 집에 반죽을 발라 집 짓기를 계속하는 꽃벌

그림 3.21. 기초공사만 마친 집에 꿀을 계속 저장하는 꽃벌
이 두 가지 실험을 보면 벌은 변화되는 상황과 관계없이 본능 속에 내재된 프로그램대로 집 짓기를 하고 꿀을 모으는 순서에 들어가며 그 순서는 바꿀 수 없다는 것을 알 수가 있다.
파브르가 행한 또 다른 실험은 벌이 꿀을 벌집에 채우는 순서와 알에서 깨어난 벌의 행동이다. 벌이 꿀을 가지고 집에 도착하면 먼저 꿀주머니 속의 꿀을 토해 내려고 벌집 속으로 머리를 넣는다. 그런 후 꿀을 토해내고 머리를 뺀 후 대신 꽁무니를 넣어서 몸에 묻어 있는 꽃가루를 털어낸다. 그런데 벌이 벌집에 꿀을 토해내고 꽁무니를 넣는 순간 꽁무니를 못 넣게 집 밖으로 쫓아내면 어떤 행동을 할까? 그럴 경우 벌은 다시 날아와 조금 전에 꿀을 토해 몸 안의 꿀주머니가 비어 있는데도 다시 머리부터 먼저 넣고 꿀을 토한 후 꽁무니를 넣는 동작을 반복한다.
이 실험 역시 벌이 본능 속에 내재된 꿀을 채우는 프로그램대로 행동하며 그 순서는 바꿀 수 없다는 것을 나타낸다.
그림 3.22. 벌은 꿀을 먼저 토해내고 꽃가루를 턴다(좌). 꽃가루를 털어내는 벌을 방해하면 다시 꿀을 토하고 꽃가루를 털려고 한다(우)
벌이 알에서 깨어나 밀봉된 집을 뚫고 나오는 행동에서도 이와 비슷한 본능을 관찰할 수 있다. 미장이 꽃벌은 집 짓기가 끝나면 꿀을 채워 넣고 그 위에 알을 낳은 후 집 입구를 막아 버린다. 막힌 입구는 시멘트처럼 딱딱하지만 알에서 깨어난 성충은 곧바로 크고 튼튼한 턱으로 그 단단한 벽을 뚫고 밖으로 나온 후 꿀을 찾으러 날아간다. 그런데 벌집 입구에 원뿔 모양의 종이 뚜껑을 씌워 놓으면 딱딱한 벽을 뚫고 나온 성충은 어떻게 행동할까? 시멘트처럼 딱딱한 벽을 뚫을 수 있는 턱을 가지고 나왔지만 그것보다 훨씬 더 뚫기 쉬운 종이 뚜껑에 부딪히게 되면 종이 뚜껑을 뚫으려는 노력을 전혀 하지 않고 그대로 굶어 죽는다.

그림 3.23. 집에 딱 붙은 종이는 뚫고 나오지만 원뿔모양의 종이뚜껑은 어떻게 해야 할지 몰라 굶어 죽는다
여기서 알 수 있는 사실은 벌은 태어나자마자 딱딱한 집 입구를 한 번만 뚫고 나가면 얼마든지 꽃을 찾아 날아갈 수 있는 공간이 펼쳐지리라 기대가 본능에 내재되어 있음을 알 수 있다. 하지만 뜻하지 않게 종이로 막힌 조그마한 공간을 만나게 되면 그다음에는 어떻게 해야 할지 몰라 그대로 굶어 죽게 된다. 이 사실 역시 벌이 태어나서 해야 할 행동이 톱니처럼 맞물려 있어 본능에 의한 순서대로 진행되지 않으면 자연계에서 생존할 수 없다는 것을 말해 주고 있다.
ii. 위버새의 집 짓기
아프리카나 서남아시아에서 발견되는 노란 위버새의 집 짓기를 살펴보면 큰미장이 꽃벌이 보인 것과 같은 유사한 본능을 발견할 수 있다. 위버새는 좀 특이한 새집을 짓는데 풀을 하나하나 물어다가 엮어서 조롱박처럼 생긴 집을 짓는다. 마라이스(Marais)는 위버새의 이런 복잡한 집 짓기 기술이 순전히 본능적인 것인지, 아니면 학습된 행동과 관련이 있는지를 알아보기 위한 흥미로운 실험을 진행했다. 새로 태어난 위버새를 우리에 가두어 키우면서 산란기에 집을 짓지 못하게 하고 알을 낳게 하였다. 그렇게 하여 태어난 새끼에게도 집 지을 풀을 제공하지 않고 바닥에 알을 낳게 하는 실험을 네 번째 세대까지 동일하게 반복하였다. 그런 후 다섯 번째 세대에 태어난 새가 자라서 산란기를 맞게 될 때 다시 집을 지을 수 있는 풀을 제공하였는데 그 새는 본능적으로 그림에 있는 둥지와 같은 집을 만들기 시작하였다. 어떻게 한 번도 본 적이 없고 어떻게 짓는지 모르는 집을 설계도를 보듯 그렇게 완벽하게 지을 수 있을까?
여기에서 우리는 컴퓨터를 만들고 그 컴퓨터가 어떻게 작동될지 기록한 펌웨어와 위버새를 창조하시고 자연계에서 어떻게 살아갈지 본능 속에 기록한 창조주의 설계가 동일한 개념임을 알 수 있다.

그림 3.24. 위버새의 둥지
iii. 앵무조개의 집 짓기
앵무조개(Nautilus)는 열대지방 바다에서 발견되는 손바닥만 한 크기의 조개이다. 앵무조개의 껍질은 완벽한 로그 나선(Logarithmic spiral)을 따르는 아름다운 껍질을 만들어 살고 있다. 어떻게 된 일일까? 누구도 앵무조개가 고등학교 수준의 수학을 배워 집을 그렇게 지었을 거라고는 생각하지는 않을 것이다. 앵무조개가 어떻게 해서 이런 멋진 껍질을 만드는지 알아보자. 먼저 앵무조개는 등 부분의 맨틀이라는 곳에서 칼슘이 풍부한 액체를 분비하여 이전에 있던 껍질보다 지름이 조금 더 큰 원형의 껍질을 만들어 붙인다. 하지만 단순히 지름이 조금 더 큰 원들을 붙여 가면 껍질이 일직선으로 나아가 나팔과 같은 모양이 되며 동그랗게 말린 껍질 모양은 되지 않는다. 여기서 앵무조개는 놀라운 기술을 발휘하여 원형의 껍질을 만들어 붙일 때 중심에서 가까운 원의 아랫부분보다 원의 윗부분에 분비물을 조금 더 분비한다. 그렇게 되면 회전 효과를 가져와 쌓이는 원들이 직진하지 않고 로그 나선 형태의 껍질이 만들어진다.

그림 3.25. 로그 나선형 패턴의 앵무조개 껍질
여기서 우리는 다음과 같은 의문을 가진다. 앵무조개는 칼슘이 풍부한 액체를 분비하여 집을 짓는데 어떻게 하여 그런 액체가 필요한지 알았을까? 분비하는 액체의 농도, 분비 속도, 액체에서 딱딱한 껍질로 경화시키는 방법 등 어떻게 그런 정교한 생화학적 과정들을 알았을까? 앵무조개의 의지인가 아니면 본능 속에 기록된 순서에 따라 유전자 발현이 조절되어 어떤 세포에서는 칼슘 액체가 분비되고 또 다른 세포에서는 분비된 액체가 경화되는 단백질이 생성되는 등 여러 과정들이 조절되는 것일까?
이에 대한 대답을 위해 3-D 프린터(=앵무조개)로 앵무조개 껍질을 만들어 본다고 가정하자. 3-D 프린터를 처음 구입하면 아무것도 하지 않는다. 앵무조개인 3-D 프린터를 작동시키기 위해서는 3-D 프린터에 쓰이는 G 프로그래밍 언어로, 어떤 순서와 과정을 거쳐 껍질을 만들지 지시하는 프로그램을 짜야 한다. 또한 앵무조개 껍질이 만들어지는 물질을 액체 상태로 잉크 카트리지에 주입하여야 된다. 그런 후 완성된 프로그램을 입력하여 3-D 프린터를 구동하면 프린터는 입력된 프로그램이 지시하는 대로 잉크의 분사 속도를 조절해 조금씩 껍질을 만들어 나간다.
이 간단한 비유에서 알 수 있듯이 3-D 프린터인 앵무조개는 자기 의지대로 껍질을 만들지 못하고 본능 속에 입력된 G 프로그래밍 언어 같은 집 짓기 프로그램이 지시하는 대로 껍질을 만들어 낸다. 이 사실은 앵무조개의 껍질이 우연에 의한 진화의 산물이 아니라 창조주께서 앵무조개의 본능과 유전자에 내장해 놓으신 집 짓기 프로그램의 산물임을 나타낸다.
i. 자연에서 발견되는 수학적 원리와 창조론
우리가 자연계나 동식물들의 자라는 형상을 자세히 들여다보면 많은 부분에서 수학적 원리를 발견할 수 있다. 자연계에 나타난 수학적 원리중 대표적인 것은 황금비, 황금각, 피보나치 수열, 로그 나선, 프랙탈 등이다. 먼저 이 수학적 원리들을 간단히 알아보도록 하자.
그림 3.26. 황금비, 황금각, 로그 나선, 프랙탈
- 황금비는 어떤 두 수의 비율이 그 합과 두 수 중 큰 수의 비율과 같게 하는 비율(a/b=(a+b)/a)로 무리수이며 근삿값은 1.618 정도가 된다.
- 황금각: 원의 긴 원호와 짧은 원호가 황금비를 이룰 때 형성되는 각으로 무리수이며 근삿값은 137.51도 정도가 된다.
- 피보나치 수열: 0 또는 1에서 시작하며 그 뒤의 모든 항은 바로 앞 두 항의 합인 수열이다(예: 0, 1, 1, 2, 3, 5, 8, …).
- 로그 나선: 극좌표에서 반지름이 각도에 따라 기하급수적으로 증가하며, 중심에서 그은 접선이 항상 일정한 각도를 이루는 나선이며 피보나치 나선과 비슷한 형상이다.
- 프랙탈: 부분이 전체와 비슷한 형태를 반복적으로 가지며, 확대해도 동일한 구조가 나타나는 자기유사성(self-similarity)을 가진 기하학적 구조를 말한다.
이제 이러한 수학적 원리들이 자연에서는 어디에서 발견될 수 있는지 알아보도록 하자.
엽서학(Phyllotaxis)은 식물의 줄기에서 잎이 나는 잎차례를 연구하는 학문이다. 엽서학에 의하면 잎은 나사의 톱니같이 나선상으로 돌아서 나는데 잎이 얼마만큼 돌아서 처음에 난 잎과 같은 방향에서 나는지 나타내는 것을 개도(divergence angle)라 한다. 예를 들어 잎이 1-2-3-4-5 순서로 나면서 두 바퀴 돌아 6번째에서 1번과 같은 방향에서 다시 잎이 나올 경우 두 바퀴인 720도를 5번 만에 도니까 개도는 720/5=144도이고 잎이 1-2-3-4-5-6-7-8 순서로 잎이 나면서 세 바퀴 돌아 9번째에 1번 잎의 방향에서 다시 잎이 시작되면 잎은 세 바퀴인 1,080도를 8번 만에 도니까 개도는 1,080/8=135도이다. 두 예처럼 잎이 2바퀴를 5번 만에 돌면 2/5 잎차례라 부르고 3바퀴를 8번 만에 돌면 3/8 잎차례라 부른다.
그런데 식물들의 잎은 아무런 순서대로 나는 게 아니라 잎차례가 1/2, 1/3, 2/5, 3/8, 5/13… 순서로 나는데 이것을 자세히 보면 분모와 분자가 모두 피보나치 수로 되어 있다는 것을 알 수 있다. 또한 분모는 분자 피보나치 수의 다음다음 순서에 나오는 피보나치 수가 된다. 잎차례의 수가 많아지면 개도는 황금각인 137.51도에 접근한다. 개도가 황금각에 접근하면 잎차례의 중복이 최소가 된다. 이렇게 잎차례의 중복이 최소가 되면 어떤 잎이든지 햇빛을 골고루 받을 수 있고 비나 이슬이 잎으로 잘 모아져서 줄기를 타고 뿌리로 내려와 그 식물의 생존에 필수적인 물을 잘 공급할 수 있는 특징을 지니게 된다.
그림 3.27. 2/5 입차례(a) 와 3/8 입차례(b)
나뭇가지나 풀잎이 자라는 순서도 자세히 보면 피보나치 수열을 따르는데 그중 대표적인 예가 톱풀이다. 아래 그림에 있는 톱풀은 처음 가지가 하나 나온 후에 2개로 되었다가 그다음은 3개, 5개, 8개, 13개의 순서로 가지가 생기며, 각 가지에 달린 잎의 개수도 피보나치 수열을 따라 1개, 1개, 2개, 3개, 5개, 8개의 순서로 달린다.
그림 3.28. 톱풀의 가지와 잎
식물의 잎뿐만 아니라 새싹, 열매, 씨앗도 피보나치 수열과 황금각의 지배를 받는다.
노르웨이 가문비나무의 싹이 트는 패턴은 피보나치 수열과 황금각의 원리를 따른다. 다음 그림에서 보이듯이 중심에서 각도를 재어보면 각각의 새싹은 이전 싹에서 약 137.5도(황금각)의 각도로 나온다. 그 결과 가지가 줄기를 중심으로 피보나치 수열의 분포와 일치하는 나선형 패턴으로 형성된다. 이 자연스러운 패턴은 나무가 햇빛, 물, 영양분을 효율적으로 모을 수 있게 하여 나무를 건강하게 성장시킨다.
그림 3.29. 노르웨이 가문비나무의 발아 패턴
데이지 꽃의 꽃잎과 씨앗은 피보나치 수열을 따라 나선형으로 정렬되어 있는데, 각 방향의 나선 수는 일반적으로 21, 34와 같은 연속적인 피보나치 수다. 또한 연속된 꽃잎이나 씨앗 사이의 개도는 대략적인 황금각을 유지하며 그 감기는 패턴은 로그 나선이 된다. 로그 나선은 자기 유사성이라는 고유한 특성이 있기 때문에 데이지 꽃잎이 자라더라도 꽃잎 모양이 변하지 않고 아름다운 모양을 그대로 유지한다.
솔방울, 콜리플라워, 로마네스코 브로콜리에서도 비슷한 패턴이 발견된다. 솔방울의 비늘은 피보나치 수에 따라 나선형으로 배열되어 있으며, 일반적으로 한 방향으로는 8개, 반대 방향으로는 13개의 나선으로 감겨있으며 각 비늘은 대략적인 황금각으로 배치되어 있다. 마찬가지로 콜리플라워의 꽃봉오리는 한 방향으로는 5개의 나선으로, 다른 방향으로는 8개의 나선으로 감겨 있고, 로마네스코 브로콜리 꽃봉오리는 한 방향으로는 13개, 다른 방향으로는 21개의 나선으로 감겨있다.
파인애플은 눈 배열에서 피보나치 수열을 찾을 수 있다. 파인애플의 눈을 따라 왼쪽 위로 올라가는 나선과 오른쪽 위로 올라가는 나선, 세로로 연결된 선을 세어보면 줄의 수가 각각 5, 8, 13 또는 8, 13, 21처럼 피보나치 수열에서 연속한 세 항이 된다. 이러한 배열은 효율적인 공간 활용과 구조적 안정성을 극대화하며, 영양분을 고르게 분배하여 균일한 성장을 돕는다.

그림 3.30. 식물에서 발견되는 피보나치 수열과 로그 나선
로그 나선을 따르는 성장 곡선은 식물뿐만 아니라 사람을 비롯한 여러 동물에게서도 찾아볼 수 있는데 사람의 귓바퀴, 귀속에 있는 달팽이관, 사람의 손가락, 해마의 꼬리, 산양의 뿔, 그리고 앵무조개를 비롯한 여러 종류의 조개와 달팽이 껍질 등이다. 이렇게 동식물들의 여러 기관이나 형상들이 로그 나선 형태로 성장해야 하는 중요한 이유가 있는데, 그것은 로그 나선으로 성장할 때만 어릴 때의 모습과 똑같은 모습을 유지한 채 자라며 그 기관의 지니고 있는 고유한 기능이 커서도 잘 유지될 수 있기 때문이다.
예를 들어 사람의 귓바퀴는 소리를 잘 모으고 귀속의 달팽이관은 저음을 충분히 들을 수 있는 세기로 증폭해 주는 최적의 기하학적 모양을 지니고 있다. 그런데 만일 이런 중요한 기관들이 자라남에 따라 그 모양이 달라진다면 어떻게 될까? 그럴 경우 어릴 때는 귀가 잘 들리지만 어른이 되면 귀가 들리지 않게 되는 난감한 상황에 부딪히게 된다. 또한 로그 나선을 따라 자라는 손가락 마디들이 어릴 때는 주먹을 자유롭게 쥘 수 있을 만큼 적당한 비율이었지만 자라면서 로그 나선 형태를 벗어난다면 주먹조차 쥐기 힘든 모양의 손을 가지게 될 것이다.

그림 3.31. 달팽이관, 귀, 해마 및 손마디 뼈
자연계에서 발견되는 또 다른 수학적 원리인 프랙탈 구조는 고사리잎, 나뭇가지, 로마네스크 브로콜리, 그리고 카울리 플라워 등에서 발견된다. 우리가 고사리잎을 자세히 살펴보면 잎의 전체적인 모양이 그보다 작은 하나하나의 잎에 복사되어 나타나 있고 또 그보다 점점 더 작은 잎들에서도 전체 모양이 반복되는 구조를 볼 수가 있다.
프랙탈 구조는 식물에서뿐 아니라 우리 몸의 순환계, 신경계, 소화기관 등에서도 발견된다. 순환계는 신선한 산소와 영양분을 몸의 모든 세포에 신속하게 공급하는 역할을 해야 한다. 그러기 위해서는 몸속의 수많은 세포까지 혈관이 분포되어야 하는데 만일 이것을 보통의 방식으로 설계한다면 우리 몸을 혈관으로만 완전히 채워야만 할 것이다. 그런데 우리의 혈관이 프랙탈 구조를 갖게 만든다면 최소한의 체적을 차지하면서도 최대한의 면적을 가지게 된다. 또한 혈관이 프랙탈 구조를 가질 경우 심장의 강력한 박동을 크고 작은 수많은 혈관으로 분산시키면서 효율적으로 완충시키는 역할을 해 줄 수 있다.
인간의 심폐기관도 프랙탈 구조로 되어 있는데 폐의 큰 가지는 작은 가지로 갈라지며 작은 가지는 그보다 점점 더 작은 가지로 갈라져 마지막으로 폐포에 이르게 된다. 이렇게 프랙탈 구조로 형성된 폐의 표면적은 테니스장만 한 크기가 되어 폐로 들어온 공기에서 최대한의 산소를 흡수하게 만든다. 소화관 내부도 영양분을 효율적으로 흡수하기 위해 미세한 주름을 가진 프랙탈 구조로 되어 있는데 그 표면적은 테니스장의 4배 정도가 된다.

그림 3.32. 프랙탈 구조를 가진 고사리와 로마네스크브로콜리
자연과 생물학적 시스템에서 발견되는 황금각, 로그 나선, 피보나치 수열, 프랙탈과 같은 수학적 원리들은 방향성없는 무작위 진화의 산물이 아니라 정확한 목적으로 생명체의 유전자에 세겨진 정교한 설계의 결과임을 나타낸다. 그렇지 않을 경우 해바라기나 고사리같은 식물들이 수학을 잘하여 황금각을 따라 씨앗을 맺고 프랙탈 구조를 따라 잎을 내었다고 믿어야 할 것이다.
"수학은 하나님께서 우주를 쓰실 때 사용하신 언어다." - 갈릴레오 갈릴레이
4. 복음으로의 초대
"주의 손가락으로 만드신 주의 하늘과 주께서 베풀어 두신 달과 별들을 내가 보오니
사람이 무엇이기에 주께서 그를 생각하시며 인자가 무엇이기에 주께서 그를 돌보시나이까
그를 하나님보다 조금 못하게 하시고 영화와 존귀로 관을 씌우셨나이다
주의 손으로 만드신 것을 다스리게 하시고 만물을 그의 발 아래 두셨으니
곧 모든 소와 양과 들짐승이며
공중의 새와 바다의 물고기와 바닷길에 다니는 것이니이다
여호와 우리 주여 주의 이름이 온 땅에 어찌 그리 아름다운지요"(시편 8:3-9)
위의 성경 구절에서 시인은 하늘과 천체를 관조하면서 그 만물을 만드신 창조자를 찬양하고 인간인 우리 자신이 누구인지를 돌아보며 읊은 시이다. 이 책에서 우리는 광대한 우주로부터 지구, 그리고 특히 지구에서 모든 생명체가 존재하는 것은 결코 우연의 결과가 아니라 창조주 하나님의 지혜와 능력으로 지어진 것들이라는 것을 분명하게 보았다. 그렇다면 하나님께서 왜 우주를 창조하셨으며 이 지구상에 생명체를 지으시고 나를 인간으로 만드셔서 살아가도록 하셨을까? 성경은 창조의 목적을 분명히 밝히고 있으며 하나님께서는 우리 인간과 영원한 교제를 원하심을 알수가 있다. 또한 성경에서는 "땅의 모든 끝이여 내게로 돌이켜 구원을 받으라"(이사야 45:22)라고 말씀하신 것처럼 하나님께서는 우리에게 구원을 주시어서 하늘나라에서 영원한 천국 시민으로 살아가기를 바라신다. 그러나 많은 사람들은 이 세상의 삶이 끝나면 모든 것이 끝나는 줄 알고 이 세상의 일에만 몰두하여 하나님께서 주신 구원의 기회를 놓치고 있다. 예수님께서 이 사실을 성경의 ‘큰 잔치 비유(누가복음 14:16-24)’와 ‘혼인 잔치 비유(마태복음 22:1-14)’에서 정확히 묘사하고 있다. 이 비유에서 사람들은 각자의 분주한 핑계를 대며 천국에로의 초대에 응하지 않으려 한다. "나는 밭을 샀으므로…", "나는 소를 시험하러 가야 하므로…", "나는 장가들었으므로…" 등으로 초대를 거절한다. 친구와 두는 바둑에서조차 몇 수 앞을 내다보는 사람들이 가장 중요한 인생에서 한 수 앞을 내다보지 않는다는 것은 안타까운 일이다.
이 장에서는 여러분을 천국으로 인도하는 복음에 대해 소개하고자 한다. 하지만, 여전히 우주와 만물을 통해 계시된 하나님의 존재를 믿기 어려워하는 사람들을 위해 하나님의 존재를 믿는 것이 합리적이라는 파스칼의 내기(Pascal’s wager)를 소개하기로 하겠다.
철학자이자, 물리학자이자, 수학자이자, 신학자인 파스칼은 그의 저서 《팡세》에서 사람들을 진리로 이끌려고 기독교의 교리를 설명하며 변증하였다. 그는 책에서 인간은 광활한 우주에 비하면 겨우 하나의 점과 같은 갈대처럼 약한 존재지만 그것은 ‘생각하는 갈대’라는 유명한 말을 남겼으며, “공간에 의해서 우주는 나를 감싸고, 하나의 점으로서 나를 삼킨다. 그러나 사고에 의해서 나는 우주를 감싼다.”라는 명언도 남겼다. 파스칼은 ‘인간의 기쁨은 창조주 신을 아는 것’이라고 하였으며 신의 존재에 대해 다음과 같은 파스칼의 내기를 제시하였다.
| 하나님께서 존재하심 | 하나님께서 존재하지 않으심 |
하나님의 존재를 믿음 | 영원한 기쁨(천국) | 바르게 세상을 삶 |
하나님의 존재를 믿지 않음 | 영원한 고통(지옥) | 무신론자로 삶 |
표 4.1. 파스칼의 내기
지금까지 우리는 천지 만물이 결코 우연에 의하여 존재하는 것이 아니라 창조주 하나님의 손길에 의해 창조되었다는 것을 살펴보았다. 따라서 우리에게 주어진 파스칼의 내기에는 두 가지 선택이 있다. 하나님의 존재를 믿고 영원한 하나님의 나라인 천국으로 들어가는 길을 택하든지 아니면 믿지 않고 영원한 멸망의 길인 지옥으로 가는 경우이다. 이 선택에 대해 누구라도 첫 번째를 선택할 것이다. 천국과 지옥은 여러분에게 신화처럼 들릴지 모르겠지만 하나님의 말씀인 성경에 의하면 반드시 존재한다.
성경 고린도후서 12장 2-4절을 보면 다음과 같은 구절이 나온다.
“내가 그리스도 안에 있는 한 사람을 아노니 십 사년 전에 그가 셋째 하늘에 이끌려 간 자라(그가 몸 안에 있었는지 몸 밖에 있었는지 나는 모르거니와 하나님은 아시느니라)
내가 이런 사람을 아노니(그가 몸 안에 있었는지 몸 밖에 있었는지 나는 모르거니와 하나님은 아시느니라)
그가 낙원으로 이끌려가서 말할 수 없는 말을 들었으니 사람이 가히 이르지 못할 말이로다.”
위 경험을 고백한 사람은 사도 바울로서 그리스도인들을 박해하기 위해 다마스쿠스로 가던 중에 예수님을 만나 극적으로 회심하게 되었고 이후에 열정적으로 예수님을 증거하는 전도자가 되었다. 그의 고백을 보면 천국 낙원은 셋째 하늘인 초공간에 위치하며 육으로는 못 가고 영으로만 갈 수 있는 것으로 생각된다. 또한, 삼차원에 살았던 바울이 이해하지 못하는 놀랍고도 신비로운 경험을 하였지만 사람들에게 말해서는 안 되는 큰 비밀이 담겨 있는 듯하다.
하나님께서는 모든 사람을 영원한 생명을 얻고 천국에서 살 수 있도록 초청하고 계신다. 이 일을 위하여 하나님께서는 영원 전부터 함께 계셨던 예수 그리스도를 이 땅에 보내셨다. 예수님이 이 땅에 오신 것은 역사적 사건이며 우리 인류의 역사는 예수님이 오시기 전 BC(Before Christ)과 오신 후 AD(Anno Domini: In the year of the Lord)로 나누어진다.
이 땅에 오신 예수님은 세상의 어떤 인간도 행할 수 없는 수많은 이적을 행하셨고 하늘에서 오신 분만이 할 수 있는 가르침을 주셨다. 죽은 나사로를 살리시거나, 날 때부터 눈먼 자와 같은 장애인이나 병자를 치유하시거나, 오병이어로 수천 명을 먹이시거나, 바람과 파도를 다스리시거나, 물 위로 걸으시는 등 여러 이적을 행하셨고, 마귀들도 그 앞에 떨며 순종하였다. 예수님은 죄로 말미암아 고통 중에 있는 죄인의 죄를 용서하시고 질병에서 치유하심으로 하나님만이 할 수 있는 죄 용서의 권세도 가지신 분임을 보여 주셨다. 이러한 이적들은 오직 생명의 주, 자연 만물의 주만이 행하실 수 있는 이적이었고 이러한 이적들과 천상의 가르침을 통하여 예수님 자신이 진정으로 하나님에게서 오신 구원자이심을 드러내어 사람들로 믿을 수 있는 분명한 증거를 주셨다.
예수님이 이 땅에 오신 궁극적인 목적은 우리를 죄에서부터 구원하시기 위해서이다. 죄란 하나님을 믿지 않고 자기중심의 삶에서 나오는 불신앙과 거짓, 사기, 우상 숭배, 탐욕, 교만, 음행, 분노, 시기, 질투 등을 말하며 누구라도 이런 죄에서 자유로운 사람은 없다. 이 죄로 인해 하나님과 인간 사이의 관계는 단절되었다. 하지만 당신의 형상대로 만들어진 인간을 너무나 사랑하시는 하나님께서는 이 무서운 죄의 문제 해결을 위해 예수 그리스도를 통하여 우리를 구원하기 위하여 찾아오셨다. 의로우신 예수 그리스도는 십자가에서 죄인들인 우리의 모든 죄짐을 지시고 죄의 대가인 죽음을 대신 치러 주셨다. 그리고 장사한 지 사흘 만에 부활하심으로 인간의 죽음을 정복하여 주셨고 제자들이 보는 가운데 하늘에 승천하셔서 영원한 주님으로 살아서 통치하고 계신다.
따라서 누구든지 자신이 하나님 앞에 자신이 죄인임을 고백하고 자신의 죄를 위해 십자가에서 대신 돌아가시고 부활하신 예수님을 구주로 영접하면 죄 용서를 받고 하나님의 영으로 말미암아 하나님의 자녀로 거듭 태어난다. 거듭난 자는 영생을 얻게 되며 하나님의 나라 백성이 된다. 이것이 복음이다. 너무 큰 은혜라 쉽게 믿기지 않지만 사실이다. 이 복음이 우리가 우리의 죄로 인하여 멀어졌던 하나님과의 관계를 올바르게 회복시킨다. 영생은 하나님께서 우리에게 주시는 은혜의 선물이기 때문에 우리 자신이 착하게 산다고만 해서는 결코 얻어질 수가 없다. 이 영생의 선물을 받기 원한다면 주일날 가까운 교회로 나가 예수님을 구주로 영접하고 싶다고 목사님께 말씀드리기를 바란다. 그러면 목사님께서 다음과 같은 영접 기도와 함께 예수님의 하신 일과 아울러 믿음의 생활에 대하여 잘 알려 주실 것이다:
하나님 아버지,
저는 죄인입니다. 지금까지 어디에서 와서 어디로 가는지도 알지 못한 채 방황하며 살았습니다. 제 욕심과 육신의 욕망을 따라 살면서 하나님께서 기뻐하시지 않는 많은 죄를 지었습니다.
하지만 하나님께서는 저를 사랑하시어 예수 그리스도를 이 땅에 보내주셨고, 예수님께서 저의 모든 죄를 대신 지시고 십자가에서 죽으셨으며, 사흘 만에 부활하셨음을 믿습니다.
이제 진심으로 제 마음을 열고 예수님을 제 구주로 모시기를 원합니다. 저를 받아 주시고, 저의 죄를 용서하시며, 영원한 생명의 길로 인도하여 주옵소서.
예수님의 이름으로 기도드립니다. 아멘.
예수님을 구주로 영접하고 나면 불안했던 마음에 천국의 평안이 임할 것으로 믿는다. 예수님이 나의 주인이신데 무엇이 두려울 것인가? 또한, 세상을 보는 눈이 달라지고 삶의 우선순위가 바뀔 것으로 확신한다. 땅 위의 것만 추구하다가 허무하게 끝나는 인생이 아니라 하늘의 영원한 것을 추구하고 나아가는 새로운 인생으로 변화될 줄 믿는다. 이번 주일이 여러분에게 이런 새로운 영적 세계를 안내하는 인생 최고의 값진 주일이 되길 기대해 본다.
“주 예수 그리스도를 믿으라. 그리하면 너와 네 집이 구원을 받으리라.”(사도행전 16:31)
추천의 글
이 책의 저자이신 김동찬 박사를 처음 만난 것은 20여 년 전 일본의 카나가와켄의 야마토시에서 개척교회인 야마토 침례교회를 담임한 지 4년 정도 되었을 때였다. 김 박사께서 일본 宇宙航空研究開発機構에 천문학 연구차 와서 주일 날 하나님께 예배하기 위하여 마침 제가 섬기는 교회에 방문한 것이었다. 예배 후에 인사하면서 김 박사께서는 그리스도인 천문학 박사라는 것을 알게 되었다.
그때까지 나는 천문학에 대하여 아무런 조예도 없었지만 천문학자의 지식과 신앙에 대하여 약간의 호기심이 들었다. 그래서 혹시 천문학자로서 교회에서 세미나를 할 수 있는지 여쭈어보았다. 개척교회라 그간 외부 강사를 모신 적도 없었기에 뭔가 새로운 집회나 세미나를 열면 성도님들도 신선한 경험을 할 기회가 될 거 같았기에 한번 가볍게 부탁을 한 것이었다. 김 박사께서는 이미 일부 교회에서 본인이 이러한 세미나를 하신 경험이 있으시다고 하셨다. 날짜를 정하였고 간단하게 만든 포스트를 시내의 몇 군데 게시판에도 붙여서 홍보도 하여 일본인들도 초청하여 천문학적 관점에서의 신앙과 창조론에 대하여 일일 세미나를 하게 되었다. 상가 건물 2층의 좁은 교회당에 우리 교회 성도님들과 몇 명의 일본인들이 참석하였지만 김 박사는 정성을 다하여 세미나를 인도하여 주었다.
이 세미나를 통하여 참석한 이들은 깊은 감명을 받았는데 특히 나에게 이 세미나는 큰 감동으로 다가왔다. 이 감동으로 나는 김 박사의 이 세미나를 좀 더 많은 사람들이 참여하여 들었으면 하는 바람이 있어 기회를 찾아보았다. 마침 카나카와켄의 침례교회 목회자들의 정례 모임에서 만나서 잘 알고 지내던 히라츠카시에 있는 침례교회의 일본인 카지이이 요시로 목사님은 매년 연말에 시민들을 대상으로 교회당이 아닌 시내의 큰 극장이나 강당을 빌려서 수많은 시민들을 초청하여 시민 크리스마스 공연을 개최하고 계셨는데 김 박사의 세미나를 소개하여 드렸다. 그해 연말에 김 박사는 기꺼이 미국에서 일본으로 와서 수많은 히라츠카 시민들 앞에서 천문학적 관점에서 창조론에 대한 세미나를 인도하여 주셨다.
김 박사의 세미나에서 나에게 온 변화는 인간으로서 나의 생명과 내 주위에 늘 당연히 존재하는 것들로 무심하게 여겨 온 것들, 무관심하게 대하여 온 것들이 얼마나 귀하고 소중한지를 새롭게 보는 눈이 열렸다는 것이다. 이것은 내가 십 대 후반 처음 주님을 만나서 내 영혼이 거듭남으로 나의 영안이 열려 만물을 새롭게 보게 된 큰 감동과는 또 다른 차원의 자연과 생명에 대한 나의 심령의 눈이 열리는 순간이었다.
이 크고 큰 우주에 흩어져 있는 무수한 은하들과 그 속의 수많은 행성들 가운데 이 지구라는 작은 행성이 특별히 선택되어 생명체가 존재하는 환경으로 창조주에 의하여 세밀하게 준비되고 보존되어 관리되고 있다는 것이 얼마나 경이로운 일인가? 이 지구 속의 모든 생명체들은 또 얼마나 소중하고 귀한 것들인가? 길을 가다 무심코 밟고 가는 풀 한 포기, 땅 위에 기어다니는 개미 한 마리도 얼마나 귀하고 소중한가? 그 속에 하나님의 형상으로 지음 받아 만물의 영장으로 살아가는 인간인 나는 누구이며 무엇이길래 이렇게 존귀한 존재로 빚어져 살아가고 있을까?
김 박사의 세미나에서 내가 받은 감동과 자각은 평생을 배우는 자로 살고 싶어 하는 내가 배우고 경험한 것들에게서 느낀 신선한 감동들이나 만족감과는 비교될 수 없는 것들이었다. 왜냐하면 그의 세미나는 단지 현대 천문학적인 연구 결과만을 소개하는 것을 넘어서서 우리 생명 자체의 근원지로 청중들을 이끌어 가서 크고 크신 하나님에 대한 경외감과 아울러 생명체의 가치와 그 숭고성을 보게 하기 때문이다.
나에게 지금까지 진한 여운으로 남>아 있는 김 박사의 세미나는 20여 년 전에 개최된 것이었고 또 제한된 시간 속에 전하여진 것이었다. 이 책에서 김 박사는 그때 세미나에서 발표한 내용들을 자세하게 설명하며 그간 연구하고 수집한 과학적 자료들을 추가로 포함하여 기술하고 있다. 또한 그가 가진 신앙으로 독자들을 창조주 하나님의 인류를 향한 영원한 구원의 계획인 복음을 받아들이도록 초청하고 있다.
아브라함 J. 헤셸은 그의 유명한 책 《사람을 찾는 하나님》의 4장 경이로움에서, 현대인들은 문명이 발전함에 따라 모든 것은 과학으로 설명이 가능하다고 생각하여 모든 사물을 당연>하게 여기는 것이 보편화되어 가므로 경이감이 사라지는 것이 이 시대정신의 한 특색이 되어 가고 있다고 지적하고 있다. 자연과 존재에 대한 경이감이 삶에 대한 감동과 감사를 불러일으킬 것이며 숭고한 경이로움을 느끼는 감정이 신앙으로 인도하는 첫걸음이 될 것이라고 하였다. 또 그는 말하기를 ‘>우리 행복의 시작은 경이로움이 없는 삶은 살 가치가 없다는 것을 이해하는 데 있으며 우리에게 부족한 것은 믿고자 하려는 의지의 부족이 아니라 경이로움을 느끼고자 하는 의지의 부족이다’라고 말하고 있다.
아무쪼록 독자 여러분들이 이 책을 통하여 제가 받은 동일한 감동으로 창조주 하나님에 대한 경외감과 그분이 창조하신 만물을 바라보는 경이로움의 눈이 열리며 여러분과 만물에 대한 하나님의 크고 놀라운 사랑을 깨닫는 기회가 되기를 바라 마지않는다.
박환철, 야마토 침례교회 목사, 일본 선교사(2000-2006)
감사의 글
이 책의 추천사를 써 주시고 책의 초고를 처음부터 끝까지 읽고 세심한 수정과 필요한 내용을 보충해 주신 캘리포니아 브리지 한인교회 박환철 선교사님에게 감사드린다. 그리고, 성경과 천문학에 대한 많은 대화를 통해 책 발간 동기를 부여하신 김용철 목사님, 김종국 목사님, 김경 선교사님, 김현아 자매님에게도 감사드린다. 또한, 원고를 읽고 피드백을 주신 샬러츠빌 블루진 한인교회 임준섭 목사님, 최경주 박사님, 박지훈 박사님, 그리고 이미지 작업에 도움을 준 큰아들 사무엘, 작은아들 다니엘에게 감사드린다.
19세기 후반, 많은 젊은 미국 선교사님들이 복음을 전하기 위해 먼 한국 땅으로 건너왔다. 그들의 헌신적인 노력으로 복음이 한국 구석구석에 전해졌고, 궁극적으로 필자에게도 전해져 예수님의 은혜로 구원을 받고 믿음의 가족이 되었다. 이 자리를 빌려 그분들의 헌신과 봉사에 진심으로 감사의 말씀을 전한다.
하나님께 모든 영광을!
그림 출처
1. 우주의 창조
Fig. 1.1: NASA/JPL, Fig. 1.2: Hubble Heritage Team, Fig. 1.3: R. Hurt/JPL-Caltech/NASA, Fig. 1.4: Hubble/NASA/ESA, Fig. 1.5: Wikipedia/R. Powell, Fig. 1.6: Wikimedia/D. Leinweber, Fig. 1.7: NASA/CXC/M. Weiss(left), NASA/D. Berry (right), Fig. 1.8: Stellarium, Fig. 1.9: Physics Forums, Fig. 1.10: NASA/JPL-Caltech (left), A. Sarangi, 2018, SSR, 214, 63 (right), Fig. 1.11: Wikimedia/ALMA (ESO/NAOJ/NRAO) (left), T. Müller (HdA/MPIA)/G. Perotti (The MINDS collaboration)/M. Benisty (right), Fig. 1.12: TASA Graphic Arts, Inc., Fig. 1.14: Jon Therkildsen, Fig. 1.15: www.neot-kedumim.org.il
2. 하나님의 걸작품 지구
Fig. 2.1: R. Narasimha, Fig. 2.3: NASA, Fig. 2.4: NASA/Goddard/Aaron Kaase, Fig. 2.6: Wikimedia, Fig. 2.7: Linda Martel, Fig. 2.8: Wikimedia, Fig. 2.9: NASA/ESA/H. Weaver & E. Smith (left), NASA/HST Comet Team (right), Fig. 2.10: Wikimedia/M. Bitton, Fig. 2.11: Wikimedia/John Garrett, Fig. 2.12: UK Foreign and Commonwealth Office, Fig. 2.13: Wikipedia, Fig. 2.16: Wikipedia/G. Taylor, Fig. 2.17: NASA/Caltech
3. 창조인가 진화인가?
Fig. 3.1: Wikipedia/Yassine Mrabet, Fig. 3.2: OpenEd/Christine Miller , Fig. 3.3: Wikipedia/LadyofHats, Fig. 3.4: Wikipedia/Messer Woland & Szczepan (left), Wikipedia/LadyofHats (right), Fig. 3.5: J.E. Duncan & S.B. Goldstein, Fig. 3.6: Wikipedia/Fiona 126, Fig. 3.7: NASA, Fig. 3.8: R. Cui, Fig. 3.9: Wikipedia/Ansgar Walk, Fig. 3.10: The Whisker Chronicles, Fig. 3.11: Encyclopedia Britanica Inc., Fig. 3.12: Wikipedia, Fig. 3.13: Wikipedia/LadyofHats, Fig. 3.14: Wikipedia/J.J. Corneveaux, Fig. 3.15: Smithsonian Institution, Fig. 3.17: NRAO/AUI/NSF (left), Wikipedia/Colby Gutierrez-Kraybill (right), Fig. 3.18: Wikipedia/MikeRun, Fig. 3.20 - Fig. 3.23: Shueisha, Inc./Obara Takuya, Fig. 3.24: Wikipedia/Pinakpani, Fig. 3.25: Wikipedia/Dicklyon, Fig. 3.26: Wikipedia/Stannered (1st img), Dicklyon (2nd img), Morn the Gom (3rd img), Eequor (4th img), Fig. 3.27: M. Kitazawa/J. Plant Res., Fig. 3.28: S.R. Rahaman, Fig. 3.30: Jill Britton (pineapple), Fig. 3.32: Wikipedia/Farry (left), Wikimedia/Ivar Leidus (right).
참고 문헌
1. 우주의 창조
제자원 (2002), Oxford Bible Encyclopedia, Bible Textbook Co., Genesis Chap. 1-11.
Another universe existed before Big Bang? 우주먼지의 현자타임즈, 2/24/2024, https://www.youtube.com/watch?v=RckLkaVzFe0
A Big Ring on The Sky: AAS 243rd Press conference. Alexia M. Lopez, 1/11/2024, https://www.youtube.com/watch?v=fwRJGaIcX6A
Bogdan, A., et al. (2024), “Evidence for heavy-seed origin of early supermassive black holes from a z ≈ 10 X-ray quasar”, Nature Astronomy, 8, 126.
Bonanno, A., & Fröhlich, H.-E. (2015), “A Bayesian estimation of the helioseismic solar age”, Astronomy & Astrophysics, 580, A130.
Karim, M. T., & Mamajek, E. E. (2017), “Revised geometric estimates of the North Galactic Pole and the sun's height above the Galactic mid-plane”, MNRAS, 465, 472.
Lopez, A. M., et al. (2022), “Giant Arc on the sky”, MNRAS, 516, 1557.
Lopez, A. M., Clowes, R. G., & Williger, G. M. (2024), “A Big Ring on the Sky”, JCAP, 07, 55.
Lyra, W., et al. (2023), “An Analytical Theory for the Growth from Planetesimals to Planets by Polydisperse Pebble Accretion”, The Astrophysical Journal, 946, 60.
Penrose, R. (2016), The Emperor’s New Mind, Oxford University Press, Oxford, United Kingdom.
Perotti, G., et al. (2023), “Water in the terrestrial planet-forming zone of the PDS 70 disk”, Nature, 620, 516.
Sandor, Zs., et al. (2024), “Planetesimal and planet formation in transient dust traps”, Astronomy & Astrophysics, in press.
Schiller, M., et al. (2020), “Iron isotope evidence for very rapid accretion and differentiation of the proto-earth”, Science Advances, 6, 7.
Tonelli, G. (2019), Genesis: The story of how everything began, Farrah, Straus and Giroux, New York, pp 19-44
Tryon, E. P. (1973), “Is the Universe a vacuum fluctuation”, Nature, 246, 396.
Vorobyov, E. I., et al. (2024), “Dust growth and pebble formation in the initial stages of protoplanetary disk evolution”, Astronomy & Astrophysics, 683, A202.
Yi, S., et al. (2001), “Toward Better Age Estimates for Stellar Populations: The Y2 Isochrones for Solar Mixture”, The Astrophysical Journal Supplement Series, 136, 417.
2. 하나님의 걸작품 지구
Comins, N. F. (1993), What If the Moon Didn't Exist? HarperCollins Publishers Inc., New York, NY.
Gonzalez, G. & Richards, J. W. (2004), The privileged planet: How Our Place in the Cosmos Is Designed for Discovery, Regnery Publishing, Inc.
Lineweaver, C. H., et al. (2004), “The Galactic Habitable Zone and the Age Distribution of Complex Life in the Milky Way”, Science, 303 (5654), 59.
Lüthi, D. et al. (2008), “High-resolution carbon dioxide concentration record 650,000 - 800,000 years before present”, Nature, 453, 379.
Narasimha, R., et al. (2023), “Making Habitable Worlds: Planets Versus Megastructures”, arXiv:2309.06562.
OpenAI. (2024), ChatGPT (4o) [Large language model], https://chatgpt.com
Ward, Peter D. & Brownlee, Donald (2000), Rare Earth: Why Complex Life is Uncommon in the Universe, Copernicus Books (Springer Verlag).
3. 창조인가 진화인가?
Abelson, P. H. (1966), “Chemical Events on the Primitive Earth”, Proc Nat Acad Sci, 55, 1365.
Behe, M. J. (2006). Darwin's black box: The biochemical challenge to evolution. Free Press.
Behe, M. J. (2020). Darwin devolves: The new science about DNA that challenges evolution. HarperOne.
Bernhardt, H. S. (2012), “The RNA world hypothesis: the worst theory of the early evolution of life (except for all the others)”, Biology Direct, 7, Article number: 23.
Chyba, C. F., & Sagan, C. (1992), “Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: An inventory for the origins of life”. Nature, 355, 125.
Cui, R., “The transcription network in skin tanning: from p53 to microphthalmia”, https://www.abcam.com/index.html?pageconfig=resource&rid=11180&pid=10026
Dembski, W. A., & Ewert, W. (2023). The design inference: Eliminating chance through small probabilities. Discovery Institute.
Danielson, M. (2020), “Simultaneous Determination of L- and D-Amino Acids in Proteins”, Foods, 9 (3), 309.
Fabre, J.-H. (2015), The Mason -Bees (Perfect Library), CreateSpace Independent Publishing Platform.
Higgins, M. (2014), “Bear evolution 101”, The Whisker Chronicles, https://thewhiskerchronicles.com/2014/01/03/bear-evolution-101/
Kasting, J. F. (1993). "Earth's Early Atmosphere." Science, 259(5097), 920.
Maslin, M. (2016), “Forty years of linking orbits to ice ages”, Nature, 540 (7632), 208.
Miller, S. L. (1953), “A Production of Amino Acids under Possible Primitive Earth Conditions”, Science, 117, 528
Mumma, M. M., et al. (1996), “Detection of Abundant Ethane and Methane, Along with Carbon Monoxide and Water, in Comet C/1996 B2 Hyakutake: Evidence for Interstellar Origin”, Science, 272 (5266), 1310.
OpenAI. (2024), ChatGPT (4o) [Large language model], https://chatgpt.com
Park, Chi Hoon (2024), “Stop codon points to GOD”, Proceedings of the 20th Anniversary KRAID Symposium
Pinto, J. P., Gladstone, G. R., & Yung, Y. L. (1980), “Photochemical Production of Formaldehyde in Earth’s Primitive Atmosphere”, Science, 210, 183.
Pinto, O. H., et al. (2022), “A Survey of CO, CO2, and H2O in Comets and Centaurs”, Planet. Sci. J., 3, 247.
Russo, D., et al. (2016), “Emerging trends and a comet taxonomy based on the volatile chemistry measured in thirty comets with high resolution infrared spectroscopy between 1997 and 2013”, Icarus, 278, 301.
Sanjuán, R., Moya, A., & Elena, S. F. (2004), “The distribution of fitness effects caused by single-nucleotide substitutions in an RNA virus”, Proc Natl Acad Sci, 101(22), 8396.
Trail, D., et al. (2011), “The oxidation state of Hadean magmas and implications for early Earth’s atmosphere”, Nature, 480, 79.
Urey, H. C. (1952). "On the Early Chemical History of the Earth and the Origin of Life." Proc Natl Acad Sci, 38(4), 351.
Wikipedia, Mutation (Distribution of fitness effects).
Wikipedia, Visual phototransduction.
Yang, P.-K. (2016), “How does Planck’s constant influence the macroscopic world?”, Eur. J. Phys., 37, 055406.
Zahnle, K. J. (1986), “Photochemistry of methane and the formation of hydrocyanic acid (HCN) in the Earth’s early atmosphere”, J. Geophys Res, 91, 2819.
About the Author
Dr. Dongchan Kim earned his B.S. in Astronomy from Yonsei University in Seoul, Korea, and his Ph.D. in Astronomy from the University of Hawai‘i at Mānoa. After completing his doctoral studies, he pursued astronomical research at several institutions, including NASA's Jet Propulsion Laboratory/Caltech, Seoul National University, and the University of Virginia.
Dr. Kim's research focuses on luminous infrared galaxies (LIRGs), ultraluminous infrared galaxies (ULIRGs), quasars, and recoiling supermassive black holes.
He is currently affiliated with the National Radio Astronomy Observatory in Charlottesville, Virginia, U.S.A.
이 책의 영어 버전은 [DIVINE GENESIS: Exploring Creation through Astronomy and Biology] 라는 제목으로 Amazon에서 출판되었다.