GÉNESIS DIVINA
Explorando la Creación a través de la Astronomía y la Biología
Introducción
Los científicos que defienden la teoría de la evolución suelen considerar que el creacionismo carece de apoyo empírico y rigor científico. Sostienen que el creacionismo no debería incluirse en los planes de estudios de ciencias, ya que no ofrece una explicación científicamente fundamentada de la diversidad y complejidad de la vida en la Tierra.
Por otra parte, la teoría evolutiva contiene lagunas y preguntas sin respuesta, sobre todo en lo que respecta al origen de la vida y la complejidad de los sistemas biológicos. La selección natural y las mutaciones son insuficientes para explicar las intrincadas estructuras y funciones observadas en los organismos vivos. Además, la teoría evolutiva sólo se aplica a los organismos vivos existentes y no aborda el origen de la vida. Además, se basa en gran medida en suposiciones y reconstrucciones especulativas, lo que pone en entredicho su validez como explicación exhaustiva de la diversidad de la vida.
Este libro está escrito para explorar el debate entre creación y evolución mediante la discusión de la creación del universo, la singularidad de la Tierra y el origen de la vida.
En la primera parte, presentaremos la estructura jerárquica del universo y analizaremos la creación del universo tal y como revelan las observaciones astronómicas. A continuación, examinaremos si la creación del universo descrita en la Biblia concuerda con los hallazgos astronómicos, si la edad de la Tierra es de 6.000 años y analizaremos más detenidamente la naturaleza de ajuste fino del universo.
La segunda parte presenta diez hechos asombrosos sobre la Tierra, destacando su idoneidad única para albergar vida y señalando pruebas de un diseño intencionado.
En la tercera parte se explora el origen de la vida, cuestionando las teorías evolucionistas convencionales y destacando la complejidad de los sistemas biológicos como prueba de la creación divina. Se examina la idoneidad del término ‘teoría de la evolución de Darwin’ y se investiga si los seres humanos evolucionaron a partir de los simios. Además, se introduce el concepto de diseño inteligente y se explora el creacionismo a través de debates sobre la física de partículas, la existencia de vida extraterrestre, los instintos de los animales y las matemáticas que se encuentran en la naturaleza.
El libro concluye con una sentida invitación a la fe, animando a los lectores a reflexionar sobre su camino espiritual y a considerar el poder transformador de la fe. Presenta el Evangelio y ofrece orientación práctica sobre cómo abrazar la fe, incluidos los pasos para comprender y recibir la vida eterna, ofreciendo esperanza y seguridad a quienes buscan una conexión más profunda con Dios.
Espero que este libro proporcione un conocimiento renovado de la creación, profundizando en la comprensión del intrincado diseño y propósito entretejidos en el universo, y ofrezca la oportunidad de meditar en la gracia, la sabiduría y el poder ilimitados de Dios, el divino Creador, que sustenta todas las cosas y nos invita a maravillarnos ante Su obra.
Contenido
1. La creación del Universo
Puede que de niño recuerde noches de acampada en el campo o en lo alto de las montañas, contemplando las innumerables estrellas que brillan en la inmensidad de lo alto, o maravillándose con las estrellas fugaces que surcan graciosamente el cielo oscuro. Estas experiencias suelen llenarnos de asombro y admiración, de una profunda apreciación de la inmensa belleza y escala del universo. En esos momentos, es posible que hayas sentido una profunda conexión con el cosmos, acompañada de un sentimiento de humildad sobre el lugar que ocupas en él. Es posible que en tu mente se hayan agitado preguntas: ¿Cuántas estrellas llenan el cielo? ¿Podría haber vida más allá de nuestro mundo? ¿Cómo empezó el universo y cómo podría acabar? ¿Quién lo creó todo? La impresionante belleza y la enigmática naturaleza del cielo nocturno despiertan la curiosidad e invitan a reflexionar sobre los orígenes del universo y nuestro propósito en él. Estos momentos de fascinación dejan una huella perdurable, inspirándonos a buscar respuestas a los mayores misterios de la vida.
En este capítulo exploraremos el origen del universo desde las perspectivas astronómica y bíblica. Compararemos ambos puntos de vista para respaldar científicamente el relato de la creación del Génesis. Además, examinaremos qué fue creado primero, si la Tierra o el Sol, si la Tierra tiene 6.000 años de antigüedad y el concepto de un universo de ajuste fino.
a. La estructura jerárquica del Universo
Para discutir el origen del universo, exploremos primero su estructura jerárquica. Empezaremos por nuestro sistema solar y seguiremos con las galaxias, las galaxias externas, los cúmulos de galaxias, los supercúmulos y los complejos de supercúmulos.
i. El Sistema Solar
El sistema solar está formado por una estrella llamada Sol, ocho planetas que la orbitan, el cinturón de asteroides entre Marte y Júpiter, el Cinturón de Kuiper, y el miembro más exterior, la Nube de Oort. Una estrella se define como un cuerpo celeste autoluminoso alimentado por fusión nuclear, mientras que un planeta es un cuerpo celeste que refleja la luz de una estrella.

Fig. 1.1. Sistema solar incluyendo el Cinturón de Kuiper y la Nube de Oort
La Tierra es el tercer planeta desde el Sol. La distancia de la Tierra a la Luna es de 384.000 km, lo que llevaría 16 días en avión a 1.000 km/h. La distancia de la Tierra al Sol es de unos 150 millones de kilómetros, o una unidad astronómica (UA), lo que llevaría 17 años en avión. La distancia a Neptuno es de 30 UA, el Cinturón de Kuiper de 30 a 50 UA y la Nube de Oort de 2.000 a 200.000 UA. A la velocidad de la luz, se tardaría 8,3 minutos en viajar de la Tierra al Sol, 4 horas a Neptuno y 9,5 meses (0,79 años-luz) en alcanzar el borde interior de la Nube de Oort. En avión, tardaría unos 850.000 años.
Los cometas pueden clasificarse en cometas de corto y largo periodo. El Cinturón de Kuiper es la fuente de los cometas de periodo corto, y la Nube de Oort, la de los cometas de periodo largo. Debido a su origen, los cometas tienen órbitas muy elípticas con grandes excentricidades. El Sol tiene 109 veces el tamaño de la Tierra, 333.000 veces su masa y un periodo de rotación de unos 25 días.
ii. El sistema estelar
Al salir de la Nube de Oort, se entra en el reino de las estrellas. La estrella más cercana a la Tierra es Próxima Centauri, que tiene un 14% del tamaño del Sol, un 12% de su masa y se encuentra a unos 4,2 años luz. Viajar hasta allí en avión llevaría aproximadamente 4,6 millones de años.
Si observas de cerca las estrellas parpadeantes en el cielo nocturno, te darás cuenta de que tienen varios colores. El color de una estrella depende de la temperatura de su superficie: las estrellas más frías aparecen rojizas, mientras que las más calientes son blanquecinas. Por ejemplo, Betelgeuse (α Ori) es roja, el Sol es amarillo y Sirio (α CMa), la estrella más brillante del cielo nocturno, es blanca azulada.
La masa de una estrella determina su tasa de fusión nuclear, que a su vez rige su luminosidad y su vida útil. Las estrellas más masivas consumen su combustible más rápidamente que las menos masivas. Las estrellas terminan su vida como enanas blancas, estrellas de neutrones o agujeros negros. Las estrellas con un núcleo de masa inferior a 1,4 masas solares se convierten en enanas blancas, las que tienen un núcleo de masa comprendida entre 1,4 y 3 masas solares se convierten en estrellas de neutrones y explotan como supernovas, y las que tienen un núcleo de masa superior a 3 masas solares se convierten en agujeros negros tras pasar por la fase de estrella de neutrones. Los restos de las explosiones de supernova pueden reciclarse para formar nuevas estrellas.

Fig. 1.2. Las estrellas presentan una gran variedad de colores
Normalmente, menos de un centenar de estrellas son visibles a simple vista en una ciudad, y unas mil en el campo en condiciones ideales. La mayoría de estas estrellas se encuentran a menos de 50 años luz de la Tierra.
iii. Nuestra Galaxia (Vía Láctea)
La Vía Láctea es una galaxia espiral barrada que contiene entre 200.000 y 400.000 millones de estrellas, además de grandes cantidades de gas, polvo y materia oscura. Su diámetro es de unos 100.000 años-luz y su grosor de unos 1.000 años-luz, lo que la convierte en una estructura relativamente plana en forma de disco con una protuberancia central.
El Sol está situado a unos 26.000 años-luz del centro galáctico, orbitándolo una vez cada 220 millones de años, período conocido como año galáctico. Nuestro sistema solar reside cerca del Espolón de Orión, un brazo menor situado entre los brazos espirales de Sagitario y Perseo. Situado a unos 60 años-luz por encima del plano galáctico, este emplazamiento ofrece una perspectiva ventajosa para observar el universo en múltiples direcciones con la mínima obstrucción del denso polvo y gas del disco galáctico.

Fig. 1.3. Nuestra galaxia (Vía Láctea)
iv. Galaxias, cúmulos de galaxias y supercúmulos
La Galaxia de Andrómeda (M31) es la más cercana a la Vía Láctea, situada en a unos 2,5 millones de años-luz de la Tierra. Es visible a simple vista desde el hemisferio norte (magnitud visual = 3,4) y tiene una forma similar a la de la Vía Láctea. La galaxia de Andrómeda se aproxima a la Vía Láctea a una velocidad de unos 110 km/s y se espera que colisione con ella dentro de unos 4.000 millones de años.
Las galaxias pueden clasificarse en tres grandes categorías morfológicas: espirales, elípticas e irregulares. Cuando dos galaxias espirales colisionan, sus interacciones gravitatorias pueden provocar una transformación drástica, que a menudo da lugar a la formación de una galaxia elíptica. Este proceso suele desarrollarse a través de etapas en las que interactúan galaxias, seguidas de una fase de galaxia infrarroja luminosa (LIRG) o galaxia infrarroja ultraluminosa (ULIRG).

Fig. 1.4. Galaxia espiral, galaxia elíptica y galaxia irregular
Si menos de 50 galaxias están unidas gravitatoriamente, se denominan ‘grupo de galaxias’, y si lo están cientos o miles, se llaman ‘cúmulos de galaxias’. Más de 40 galaxias cercanas, entre ellas la Vía Láctea y Andrómeda, pertenecen al Grupo Local. El Grupo Local y el Cúmulo de Virgo forman parte del Supercúmulo de Virgo, que a su vez forma parte del Supercúmulo de Laniakea.
Un complejo de supercúmulos, también conocido como filamento galáctico o cadena de supercúmulos, es una inmensa estructura a gran escala en el universo, compuesta por numerosos supercúmulos de galaxias que están interconectados por vastas redes de galaxias, gas y materia oscura. Estas regiones interconectadas forman un patrón similar a una telaraña y representan las estructuras más grandes que se conocen en el cosmos. Abarcan distancias increíbles, desde cientos de millones hasta miles de millones de años luz, empequeñeciendo estructuras cósmicas más pequeñas. Entre ellas, la Gran Muralla Hércules-Corona Borealis destaca como el mayor complejo de supercúmulos conocido, un testimonio sobrecogedor de la escala del universo. Se calcula que en el universo observable hay unos 200.000 millones de galaxias, repartidas a lo largo de una asombrosa distancia de aproximadamente 93.000 millones de años luz, cada una de las cuales contribuye a el intrincado tapiz de estructuras cósmicas.

Fig. 1.5. Supercúmulos cercanos (amarillo: supercúmulo Laniakea)
b. Creación del Universo
¿Cómo empezó el universo? ¿Ha existido siempre, o fue creado por Dios? Para explorar este tema, examinaremos el origen del universo tal como se observa en astronomía y como se describe en el Libro del Génesis de la Biblia.
i. Creación del Universo en Astronomía
La teoría más extendida sobre el origen del universo es la del Big Bang, que postula que el universo comenzó hace aproximadamente 13.800 millones de años como un punto increíblemente caliente y denso que se expandió rápidamente. Esto plantea naturalmente la intrigante pregunta: ‘¿Qué existía antes del Big Bang?’ Una de las principales hipótesis afirma, con creciente apoyo en, que antes del Big Bang el universo existía en un estado de fluctuaciones cuánticas dentro del vacío, una base dinámica y probabilística de la que surgió nuestro universo.
Antes de Paul Dirac, el vacío se consideraba un espacio vacío sin nada. En 1928, Dirac combinó la mecánica cuántica y la relatividad especial para describir el comportamiento de un electrón a velocidades relativistas. Curiosamente, la ecuación sugería dos soluciones para el electrón: una para un electrón con energía positiva y otra para un electrón con energía negativa. Dirac propuso que el vacío no es un espacio vacío, sino que está lleno de un número infinito de electrones con energía negativa (positrón). Por ello, el vacío recibe a veces el nombre de Mar de Dirac.
Aunque el Mar de Dirac parece estático, nunca lo es debido al principio de incertidumbre de Heisenberg. Los pares de partículas y antipartículas aparecen espontáneamente (producción de pares) y desaparecen (aniquilación de pares) de forma aleatoria. La escala temporal es de 10-21 segundos e invisible para el ojo humano, pero si hay una cámara que pueda captarlo, será como mirar un mar fluctuante. Es lo que se llama ‘fluctuación cuántica’. El Big Bang surgió del mar de la fluctuación cuántica en un punto singular. El Big Bang en sí es el principio del universo.

Fig. 1.6. Modelo 3D de fluctuaciones cuánticas en el vacío
Inmediatamente después del Big Bang, el universo sufrió rápidos cambios debido a su temperatura y densidad extremadamente altas. De 10-43 segundos (tiempo de Planck) a 10-36 segundos, el universo se rigió por la Teoría de la Gran Unificación, en la que se unifican las tres fuerzas (fuerte, débil y electromagnética) del Modelo Estándar. El universo pasó por una época inflacionaria de 10-36 segundos a 10-32 segundos, una época electrodébil de 10-32 segundos a 10-12 segundos, una época de los quarks de 10-12 segundos a 10-6 segundos, una época hadrónica de 10-6 segundos a 1 segundo y una época leptónica de 1 segundo a 10 segundos.
Al final de la época leptónica se produjo un acontecimiento dramático y crucial. Los pares de leptones y antileptones, formados principalmente por electrones y positrones, se aniquilaron mutuamente. Este proceso liberó un inmenso número de fotones (partículas de luz), inundando el universo de luz. Estos fotones se convirtieron en la forma de energía dominante en el cosmos, marcando el comienzo de lo que se conoce como la época de los fotones. Esta época, que duró desde unos 10 segundos hasta 380.000 años después del Big Bang, se caracterizó por un plasma caliente y denso de electrones, núcleos y fotones libres. Durante esta época, los fotones eran dispersados por electrones y protones libres, lo que les impedía viajar libremente y hacía que el universo fuera opaco.
La época de recombinación siguió al final de la época de los fotones, en la que se produjo otro acontecimiento importante. Los electrones se combinan con protones para formar hidrógeno neutro y helio. Este es el comienzo de la era dominada por la materia. Cuando esto ocurrió, el universo lleno de plasma se volvió gradualmente transparente y se transformó en el espacio que podemos llamar cielo. Cuando esto ocurre, los fotones producidos durante la época de los fotones, pero previamente confinados por el plasma, pueden ahora moverse libremente por el universo transparente. Estos fotones que se mueven libremente se observan como luz muy brillante y forman la radiación cósmica de fondo de microondas.
Las estrellas y galaxias que vemos hoy se formaron a partir de los átomos creados durante la época de recombinación. Desde entonces, el universo ha seguido expandiéndose como consecuencia del Big Bang. Cuando el universo tenía 9.800 millones de años, la energía oscura comenzó a dominar, marcando el inicio de la era dominada por la energía oscura. En esta era, el universo continúa expandiéndose a un ritmo acelerado. Esta expansión acelerada es el estado actual del universo.
ii. El destino del Universo (¿Otra vez el Big Bang?)
El destino del universo depende de su densidad global. Según las mediciones del WMAP, la densidad actual del universo es aproximadamente igual a la densidad crítica (unos 10-29 g cm-3) con un margen de error del 0,5%. Sin embargo, esta incertidumbre significa que aún no podemos determinar definitivamente el destino final del universo hasta que se obtengan mediciones más precisas. Si la densidad del universo es superior a la densidad crítica, las fuerzas gravitatorias acabarán por superar la expansión, provocando el colapso del universo sobre sí mismo en un acontecimiento catastrófico conocido como Big Crunch, característico de un universo cerrado.
Por el contrario, si la densidad es inferior a la densidad crítica, el universo seguirá expandiéndose eternamente a un ritmo acelerado, lo que llevaría a un escenario conocido como Big Rip, característico de un universo abierto. En este caso, la temperatura del universo se enfriará progresivamente a medida que avance la expansión, y la formación de estrellas acabará por cesar debido al agotamiento del medio interestelar necesario para la creación estelar. Con el tiempo, el universo se volverá cada vez más oscuro y frío, un proceso a menudo denominado ‘muerte por calor’.
Las estrellas existentes se quedarán sin combustible y dejarán de brillar. Posteriormente, se producirá la desintegración de protones, tal como predice la Gran Teoría Unificada, cuando la edad del universo sea de unos 1032 años. Alrededor de 1043 años, los agujeros negros comenzarán a evaporarse a través de la radiación Hawking. Después de que todas las materias bariónicas hayan decaído y todos los agujeros negros se hayan evaporado, el universo se llenará de radiación. La temperatura del universo se enfriará hasta el cero absoluto y todo estará oscuro y vacío, lo que se asemeja al estado del universo sometido a fluctuaciones cuánticas antes del Big Bang.

Fig. 1.7. Destino del universo y agujero negro en evaporación
Recientemente se han descubierto dos megaestructuras cósmicas a 7.000 millones de años luz de la Tierra, en dirección a la Osa Mayor. El Arco Gigante, descubierto en 2022, y el Gran Anillo, descubierto en 2024, desafían el principio cosmológico que afirma que el universo es homogéneo e isótropo a gran escala. Estas megaestructuras requieren una explicación adecuada. Una posible explicación es que sean enormes cuerdas cósmicas o restos de la evaporación Hawking de agujeros negros supermasivos (puntos Hawking) del Big Bang anterior.
Esta interpretación está relacionada con la Cosmología Cíclica Conforme (CCC) de Roger Penrose. La CCC es un modelo cosmológico basado en la relatividad general, en el que el universo se expande eternamente hasta que toda la materia decae y abandona los agujeros negros. En la CCC, el universo itera a través de ciclos infinitos, con un nuevo Big Bang emergiendo dentro del Big Bang actual en constante expansion.

Fig. 1.8. Gran Anillo (azul) y Gran Arco (rojo)
Personalmente, encuentro atractivo el CCC porque ofrece soluciones potenciales a algunos problemas de la evolución de las galaxias. Existe una correlación entre la masa de un agujero negro y la dispersión de la velocidad estelar (la relación M-sigma). Según esta relación, la masa de un agujero negro es aproximadamente el 0,1% de la masa de su galaxia. Recientemente, Chandra y JWST descubrieron una galaxia intrigante, UHZ1, a través de lentes gravitacionales. UHZ1 se encuentra a una distancia de 13.200 millones de años-luz, observada cuando nuestro universo tenía sólo un 3% de su edad actual. La masa estimada del agujero negro de UHZ1 resultó ser mayor que la de la galaxia que lo alberga. Esta gran masa del agujero negro no puede explicarse con las teorías actuales sobre la masa de los agujeros negros, pero sí con la CCC. Esto puede entenderse si el agujero negro de UHZ1 era un agujero negro reciclado del Big Bang anterior y se convirtió en un agujero negro semilla en UHZ1 durante el Big Bang actual
No sabemos cómo se produce el nuevo Big Bang mientras el Big Bang actual sigue expandiéndose. Podríamos intentar utilizar el concepto de hiperespacio. En este escenario, el universo se expande en un espacio tridimensional. Sin embargo, imaginemos nuestro universo tridimensional como una superficie incrustada en un espacio de mayor dimensión (hiperespacio). Este espacio de mayor dimensión podría ser un espacio de cuatro dimensiones (o más) donde todo nuestro universo es sólo una ‘rebanada’ o una ‘brana’.
A medida que nuestro universo siga expandiéndose, podría llegar a converger en un punto singular de este hiperespacio de dimensiones superiores, de forma muy parecida a como una superficie bidimensional puede curvarse y converger en un punto del espacio tridimensional. Este punto del hiperespacio podría ser análogo al cuello de una botella de Klein, una forma de dimensión superior en la que la superficie hace un bucle sobre sí misma.
Cuando la expansión del universo en el espacio tridimensional converja hacia este punto singular en el hiperespacio, podrían crearse condiciones en las que la densidad de energía se vuelva extremadamente alta. Si este punto singular en el hiperespacio no puede acomodar la inmensa afluencia de energía y energía del vacío del universo en expansión actual, podría producirse una explosión. Esta explosión sería el comienzo de un nuevo Big Bang, que crearía un nuevo universo.
De este modo, el universo actual del Big Bang, en constante expansión, podría conducir a la formación de un nuevo universo en el marco del hiperespacio, con la convergencia a un punto singular que actuaría como puente entre los ciclos del CCC. Esta convergencia de dimensiones superiores proporciona un mecanismo para los ciclos continuos del Big Bang mientras el universo actual sigue expandiéndose, y la energía de este universo en expansión también podría contribuir a la energía oscura que impulsa su aceleración.

Fig. 1.9. Cosmología cíclica conforme
iii. La Creación del Universo en la Biblia
En esta sección analizaré la creación del universo tal como se describe en la Biblia desde una perspectiva astronómica, examinando cómo podría alinearse el relato bíblico con la comprensión científica moderna. Este análisis ahondará en los posibles paralelismos entre el relato bíblico y las observaciones astronómicas. Aunque este enfoque ofrece una perspectiva interesante, es importante reconocer que hay otras formas de interpretar el relato bíblico de la creación. Estas interpretaciones pueden variar en función de los contextos teológico, filosófico y cultural, y cada una de ellas aporta una visión única de la profunda narración de los orígenes del universo.
a) Dios declaró la creación del universo
La creación del universo se describe en el Génesis, el primer libro de la Biblia.
“En el principio creó Dios los cielos y la tierra.” (Génesis 1:1)
Este versículo introduce el acto de la creación por Dios, afirmando que Él es el iniciador de todo lo que existe. La frase “los cielos y la tierra” abarca toda la creación, indicando la totalidad del universo.
“Y la tierra estaba desordenada y vacía, y las tinieblas estaban sobre la faz del abismo, y el Espíritu de Dios se movía sobre la faz de las aguas.” (Génesis 1:2)
El término “tierra” representa aquí la creación física, material (es decir, la materia bariónica) a la que Dios daría forma más tarde. La frase “la tierra estaba desordenada” puede interpretarse como la descripción de un estado primordial de vacío, en el que aún no se había creado nada. El término “vacía” significa un espacio vacío, y si no hay nada dentro de ese espacio, puede llamarse legítimamente vacío. Por lo tanto, la frase “la tierra estaba desordenada y vacía” sugiere que, desde el principio, el universo existía como un vacío, un estado inicial de nada. La siguiente frase “las tinieblas estaban sobre la faz del abismo” tiene un significado profundo. La “tinieblas” es חֹשֶׁך (choshek) en hebreo y significa literalmente oscuridad total sin ninguna luz. Lo “abismo” es תְּהוֹם (tehom) en hebreo y se derivó de הום (hom) que significa 'alboroto' o 'fluctuar'. Por lo tanto, “Y la tierra estaba desordenada y vacía, y las tinieblas estaban sobre la faz del abismo” puede interpretarse como una descripción del origen del universo a partir de un vacío en un estado de oscuridad y fluctuación. Esta interpretación concuerda estrechamente con la condición del universo en su etapa más temprana, justo antes del Big Bang, cuando existía como un vacío sometido a fluctuaciones cuánticas.
b) La creación de la luz
El acontecimiento principal del primer día de la creación es la creación de la luz.
“Y dijo Dios: Sea la luz; y fue la luz.” (Génesis 1:3)
El versículo afirma que Dios inició la creación del universo creando la luz. De manera similar, el Big Bang comenzó con una serie de épocas rápidas, que en conjunto duraron menos de un segundo, y que finalmente condujeron a la creación de la luz (fotones) durante la época de los fotones. La creación de la luz en Génesis 1:3 se corresponde de manera notable con la creación de la luz durante la era de los fotones, lo que alinea poderosamente el relato bíblico con este momento crucial en los inicios del universo.
c) La creación del cielo
El acontecimiento principal del segundo día de la creación es la creación del cielo.
“E hizo Dios la expansion, y..., Y llamó Dios a la expansión Cielos...”. (Génesis 1:7, 8)
La creación del cielo descrita en el Génesis puede correlacionarse con la época de recombinación de la cosmología del Big Bang. Antes de esta época, el universo era opaco, lleno de un plasma denso y caliente de electrones, neutrones, protones y fotones. Este plasma dispersaba los fotones, impidiéndoles viajar libremente y haciendo que el universo fuera opaco a la radiación. Durante esta época, el universo tenía unos 10 años-luz de diámetro, lo que significaba que no había espacio despejado para un ‘cielo’ visible.
Sin embargo, en la época de recombinación, el universo se enfrió lo suficiente como para que electrones y protones se combinaran y formaran átomos neutros de hidrógeno. Este proceso limpió el plasma, haciendo el universo transparente y permitiendo que los fotones viajaran libremente por el espacio. Como resultado, surgió una vasta extensión transparente, lo que conocemos como el cielo visible, con un radio de unos 42 millones de años-luz. Por lo tanto, la creación del cielo en Génesis 1:7-8 puede interpretarse como una referencia a este acontecimiento crucial en la historia cósmica.
La siguiente tabla resume la creación del universo tal y como se describe en la Biblia y como la explica la astronomía. La comparación muestra que el relato de la creación en el Génesis concuerda con los hechos astronómicos en un grado notable, lo que confirma que Dios ya había revelado estas verdades a través de la Biblia mucho antes de que fueran descubiertas por la ciencia.
Génesis | Astronomía |
Fluctuación del vacío (Gn 1:2 - antes de la Creación) | Fluctuación del vacío (antes del Big Bang) |
Creación de luz (Gn 1:3 - Día 1 de la Creación) | Creación de luz (Época fotónica) |
Creación del cielo (Gn 1:7-8 - Día 2 de la Creación) | Creación del cielo (Época de recombinación) |
Tab. 1.1. Comparación de la Creación en el Génesis y la Astronomía
c. ¿Qué se creó primero, la Tierra o el Sol?
El acontecimiento principal del tercer día de la creación en el Génesis es la creación de la tierra seca y el mar. Puede entenderse como el periodo durante el cual se formó y estructuró la Tierra. El proceso de acumulación de agua y revelación de la tierra seca significa el desarrollo de la superficie terrestre y de los accidentes geográficos. El acontecimiento principal del cuarto día del Génesis es la creación del Sol. Así pues, la Tierra fue creada antes que el Sol. Será interesante examinar si el relato bíblico es coherente con las observaciones astronómicas. Explorémoslo.
Las estrellas y los planetas se forman a partir de nubes moleculares. Las nubes moleculares están formadas por un 98% de gas (un 70% de hidrógeno y un 28% de helio) y un 2% de polvo (carbono, nitrógeno, oxígeno, hierro, etc.). La mayoría de las estrellas y los planetas jovianos están formados por gas, y la mayoría de los planetas terrestres, por polvo. Las protoestrellas se forman cuando las nubes moleculares colapsan bajo su propia gravedad. Durante este proceso, el material restante de las nubes moleculares forma un disco giratorio conocido como disco protoplanetario, que es la región donde los planetas acaban tomando forma. El colapso gravitatorio inicia el calentamiento y la compresión del núcleo, dando lugar al nacimiento de una protoestrella, mientras que el disco giratorio circundante proporciona el entorno para la formación y evolución de los cuerpos planetarios.

Fig. 1.10. Protoestrella y disco protoplanetario, y diagrama H-R
A medida que la protoestrella continúa contrayéndose, se convierte en una estrella de la pre-secuencia principal y sigue las vías de evolución estelar conocidas como la vía de Hayashi (para estrellas de baja masa) y la vía de Henyey (para estrellas de alta masa) en el diagrama de Hertzsprung-Russell (diagrama H-R). Las estrellas de la pre-secuencia principal pueden observarse como estrellas T Tauri si su masa es inferior a 2 masas solares, y como estrellas Herbig Ae/Be si su masa es superior a 2 masas solares. La de la pre-secuencia principal continúa contrayéndose hasta que su temperatura interna aumenta de 10 a 20 millones de grados. En este punto, la estrella de pre-secuencia principal estrella comienza la fusión nuclear del hidrógeno y se convierte en una verdadera estrella en el cielo. Las estrellas que se encuentran en esta fase se denominan estrellas de secuencia principal.
Según la teoría de la evolución estelar y los estudios de heliosismología, el Sol permaneció en la etapa previa a la secuencia principal entre 40 y 50 millones de años, tras lo cual se convirtió en una estrella de secuencia principal.
Mientras la estrella se forma en el centro, los planetas se forman en el disco protoplanetario. Las colisiones de partículas de polvo y gas forman guijarros, los guijarros se convierten en rocas y las rocas en planetesimales. Estos planetesimales son los componentes básicos de los planetas
Sólo recientemente se han estudiado activamente los detalles del proceso de formación de planetas en el disco protoplanetario. Los estudios predicen que se tardarán unos pocos millones de años en formar un planeta del tamaño de la Tierra a partir de guijarros de 1 mm de tamaño. Esta predicción puede comprobarse con observaciones reales, como las imágenes submilimétricas de ALMA de las estrellas T Tauri HL Tau y PDS 70.
La masa de HL Tau es de aproximadamente dos masas solares y su edad ronda el millón de años. La imagen revela que ya se han formado varios planetas que orbitan alrededor de la estrella central de la secuencia preprincipal, como indican los huecos en el disco protoplanetario. La masa de PDS 70 es de aproximadamente 0,76 masas solares y su edad es de unos 5,4 millones de años. Dos exoplanetas, PDS 70b y PDS 70c, han sido observados directamente por el VLT de ESO. En 2023, las observaciones espectroscópicas del telescopio espacial James Webb detectaron agua en la región de formación de planetas terrestres del disco protoplanetario y sugirieron que en su interior se han formado dos o más planetas terrestres. Es importante señalar que las nubes de gas y polvo observadas en HL Tau se eliminaron en gran medida en PDS 70, y en el centro se han formado planetas terrestres que contienen agua.
Los planetas terrestres tardaron 5,4 millones de años en formarse, pero aunque tardaran 10 millones de años, seguirían siendo muchos menos que los 40 a 50 millones de años que tardó el Sol en convertirse en una estrella de la secuencia principal. Esto sugiere que la Tierra fue creada antes que el Sol, como se afirma en el Génesis, y es coherente con las observaciones astronómicas.

Fig. 1.11. HL Tau y PDS 70
Otro acontecimiento importante que Dios realizó en el tercer día fue la creación de plantas y árboles. Los ateos y evolucionistas se preguntan a menudo cómo pudieron sobrevivir estas plantas y árboles si el Sol fue creado en el cuarto día. Esta pregunta puede abordarse en el contexto de la teoría de la evolución estelar. Cuando se formó la Tierra, el Sol aún se encontraba en la fase de estrella T Tauri. Aunque las estrellas T Tauri no son estrellas de la secuencia principal, su temperatura superficial oscila entre 4.000 y 5.000 Kelvin. La radiación de cuerpo negro a esta temperatura alcanza su punto máximo en la longitud de onda visible. Además, el tamaño del Sol cuando era una estrella T Tauri era varias veces mayor que su tamaño actual. Por lo tanto, podría proporcionar suficiente energía en el rango de longitud de onda visible para permitir la fotosíntesis en plantas y árboles.
d. ¿Tiene la Tierra 6.000 años?
El ' creacionismo de la Tierra joven ' es la creencia de que la Tierra y el universo son relativamente jóvenes, normalmente de unos 6.000 a 10.000 años de antigüedad, basada en una interpretación literal del relato bíblico de la creación en el Génesis. Los creacionistas de la Tierra joven creen que la Tierra se creó en seis días de 24- horas y rechazan gran parte del consenso científico moderno sobre la edad de la Tierra y el universo. Numerosas pruebas científicas procedentes de diversos campos, como la geología, la astronomía y la física, indican que la Tierra tiene aproximadamente 4.600 millones de años y el universo unos 13.800 millones de años. A pesar de estas abundantes pruebas, los creacionistas de la Tierra joven no están de acuerdo. Esta situación recuerda al debate entre los modelos geocéntrico y heliocéntrico en tiempos de Galileo Galilei.
Antes de entrar en el debate principal, veamos algunos ejemplos que facilitan la comprensión de que la Tierra y el universo tienen al menos varios millones de años.
La corteza terrestre está formada por placas tectónicas que se mueven lentamente, provocando terremotos. Nadie negaría este hecho. Un punto caliente es un punto en el que el magma fluye desde las profundidades del manto bajo la corteza, con su centro fijo. Cuando el magma fluye hacia la corteza y se enfría, forma tierra. Las islas hawaianas son un buen ejemplo de este proceso. En la Gran Isla de Hawai, el Kilauea sigue siendo un volcán activo, y a medida que el magma que erupciona se enfría en el agua del mar, se forma nueva tierra. La tierra recién formada se desplaza hacia el noroeste a un ritmo de unos 7-10 cm al año debido a la tectónica de placas, y este proceso ha creado las distintas islas de Hawai. Esto sucede incluso ahora, y es un hecho innegable.
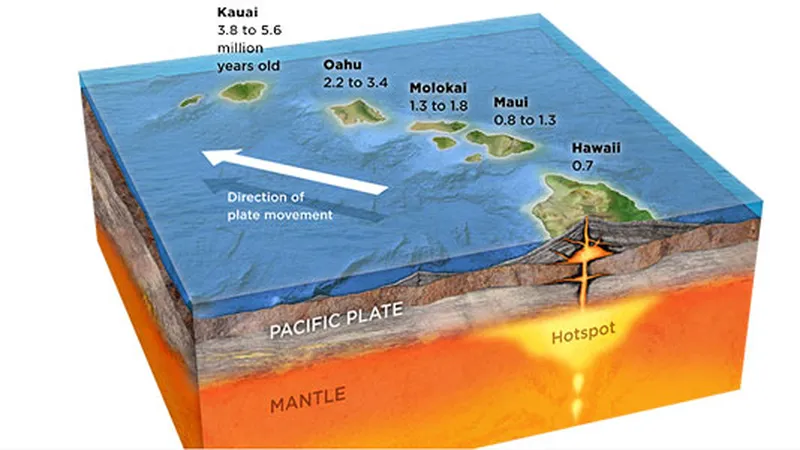
Fig. 1.12. Historia geológica de las islas hawaianas
Teniendo en cuenta la velocidad a la que se mueven las placas tectónicas, las edades de las islas hawaianas se estiman así: la Isla Grande tiene 400.000 años, Maui 1 millón de años, Molokai 1,5-2 millones de años, Oahu (donde está Waikiki) 3-4 millones de años y Kauai unos 5 millones de años. En la Gran Isla, se puede ver que gran parte de la tierra sigue cubierta de suelo volcánico negro, lo que indica una meteorización mínima. En cambio, Kauai ha sufrido una importante meteorización, lo que ha permitido que florezca la vegetación, lo que le ha valido el sobrenombre de ‘La Isla Jardín’. Este ejemplo proporciona pruebas directas de que la Tierra tiene al menos varios millones de años.
Para comprender directamente que el universo tiene al menos varios millones de años, basta con aceptar que la luz viaja a 300.000 km por segundo. El Sol está a 150 millones de km de la Tierra. Por tanto, la luz solar que recibimos ahora se generó en el Sol hace 8,3 minutos. El Sol es unas 400 veces más grande que la Luna, pero como está mucho más lejos, en el cielo parece tener aproximadamente el mismo tamaño que la Luna. Nadie lo negaría. La galaxia de Andrómeda tiene un tamaño similar al de nuestra Vía Láctea, pero se encuentra a 2,5 millones de años luz, lo que la hace parecer unas cuatro veces mayor que la Luna. El hecho de que podamos ver la Galaxia de Andrómeda significa que la luz que observamos se creó en Andrómeda hace 2,5 millones de años y acaba de llegar hasta nosotros. Si has visto la Galaxia de Andrómeda, no puedes negar este hecho. Se trata de una prueba directa de que el universo tiene al menos varios millones de años.
A pesar de estos hechos, si se sigue insistiendo en que la Tierra tiene 6.000 años, podría convertirse en un escollo más que en una ayuda para difundir el Evangelio, lo que podría distanciar a muchas personas de él. Por tanto, en lugar de defender el creacionismo de la Tierra joven, sería más razonable leer detenidamente el Génesis en la Biblia y tratar de encontrar una solución.
Para los humanos, el tiempo siempre fluye del presente al futuro y nunca fluye hacia atrás. Definimos un día como 24 horas, pero si fuéramos creados en otros planetas, un día no tendría 24 horas. Por ejemplo, si fuéramos creados en Venus, un día serían 243 días terrestres, y en Júpiter, un día serían 10 horas terrestres. Por lo tanto, a menos que cambiemos nuestra definición y percepción del tiempo desde una perspectiva geocéntrica, será difícil abordar esta cuestión. Discutámoslo más a fondo teniendo en cuenta estos hechos.
i. Los días en el Génesis
En primer lugar, estimemos la edad del universo basándonos en los registros del Génesis. Según el Génesis, Dios creó el universo y todo lo que hay en él en seis días. El tiempo transcurrido desde Adán hasta Noé puede estimarse utilizando los registros genealógicos de Génesis 5:3-32. El diluvio de Noé ocurrió cuando Noé tenía 600 años, y el número total de años desde Adán hasta el diluvio es de 1.656 años. No sabemos cuándo ocurrió el diluvio de Noé. Algunos eruditos y tradiciones bíblicas intentan datar el diluvio utilizando genealogías en la Biblia, estimando que ocurrió alrededor de 2300-2400 a.C.. Por tanto, la edad del universo, según esta interpretación, es de 7 días + 1.656 años + 4.400 años = 6.056 años. Esta es la base teórica de la afirmación de los creacionistas de que la Tierra tiene 6.000 años.
Para abordar el problema de la edad diurna, echemos otro vistazo al Génesis. Aunque no parece haber problemas con los registros genealógicos del Génesis, puede existir cierto debate sobre el año exacto del diluvio de Noé. Sin embargo, que el diluvio de Noé ocurriera hace 4.400 o 44.000 años no afecta significativamente a la edad del universo, tal como se entiende en el contexto científico de 13.800 millones de años. Entonces, ¿dónde está la clave para resolver el problema de la edad diurna? Quizá ya se haya dado cuenta: la clave está en la interpretación de los siete primeros días de la creación.
Fig. 1.13. Para definir un día, la Tierra y el Sol deben existir de antemano.
La razón es sencilla: un día se define como el periodo de rotación del planeta en el que vivimos. Para definir un día, tanto el Sol como la Tierra deben existir de antemano. Sin embargo, en Génesis se dice que la Tierra fue creada al tercer día y el Sol al cuarto, pero Dios utilizó los términos ‘día’ y ‘noche’ incluso antes de su creación. Esto implica que el ‘día’ en Génesis no es el día de 24 horas como lo definimos nosotros, sino un ‘día’ como lo definió Dios. La falacia de los creacionistas de la Tierra joven radica en su malentendido de que el ‘día’ mencionado en el Génesis se refiere a un día humano literal de 24 horas, lo que lleva a una interpretación errónea del término ‘día’ en el relato del Génesis.
Si los días del Génesis no son los periodos de 24 horas definidos por los humanos, cabe preguntarse: ‘¿Cuánto duran los días del Génesis en términos de días humanos?’. Aunque no conocemos la respuesta exacta, podemos estimar un periodo aproximado comparando los acontecimientos de la creación descritos en el Génesis con los del Big Bang.
El acontecimiento principal del primer día de la creación es la creación de la luz. La época de los fotones en el Big Bang corresponde a este acontecimiento, y el tiempo humano del primer día es de 380.000 años. El acontecimiento principal del segundo día de la creación es la creación del cielo. La época de recombinación corresponde a este acontecimiento, siendo el tiempo humano del segundo día de 100.000 años. El acontecimiento principal del tercer día es la creación de la Tierra. Como vimos en la sección anterior, la Tierra tarda unos 10 millones de años en formarse, por lo que el tercer día de la creación habría durado más de 10 millones de años. De igual manera, el evento principal del cuarto día es la creación del Sol. Dado que el Sol tarda aproximadamente entre 40 y 50 millones de años en formarse, el cuarto día de la creación habría durado más de 40 millones de años. La siguiente tabla resume los resultados anteriores.
Aquí observamos algunos hechos inesperados sobre el concepto de tiempo utilizado por Dios. Los días del relato de la creación son mucho más largos que un día humano de 24 horas. Además, el tiempo de Dios no es fijo, sino variable, desde cientos de miles de años hasta más de 40 millones de años. ¿Cómo podemos entender esto? En cierto sentido, no es un resultado sorprendente, sino esperado.
Día de la Creación | Acontecimiento en el Génesis | Evento en Astronomía | Humano tiempo |
Día 1 | Creación de la luz | Creación de la luz en la época de los fotones | 380.000 años |
Día 2 | Creación del cielo | Creación del cielo en la época de recombinación | 100.000 años |
Día 3 | Creación de la Tierra | Creación de la Tierra | > 10 millones de años |
Día 4 | Creación del Sol | Creación del Sol | > 40 millones de años |
Tab. 1.2. Días de Creación en Tiempo Humano
ii. El Creador del Tiempo
El ‘día’ utilizado en el Génesis es yom (יום) en hebreo. Yom puede interpretarse de varias maneras, entre ellas una que se refiere a la edad o a un largo periodo de tiempo. Esta interpretación sugiere que cada ‘día’ de la creación representa una larga época durante la cual tuvieron lugar actos específicos de la creación. Otra interpretación es que ‘yom’ significa un periodo de duración indeterminada. Este punto de vista postula que los días de Dios no están sujetos a las limitaciones temporales humanas, reconociendo que Dios, como creador del tiempo, opera fuera de nuestras limitaciones temporales. En la Biblia hay ejemplos de esta interpretación.
En 2 Pedro en el Nuevo Testamento, está escrito:
“Mas, oh amados, no ignoréis esto: que para con el Señor un día es como mil años, y mil años como un día.” (2 Pedro 3:8)
Este pasaje pretende animar a los que esperan las promesas de Dios a hacerlo con paciencia. También puede sugerir que la perspectiva de Dios sobre el tiempo difiere de la de los humanos, dando a entender que Dios puede dilatar o contraer el tiempo a Su antojo. Entendemos que el tiempo no es una cantidad fija. Según la relatividad especial, el tiempo transcurre más lentamente para el observador en movimiento que para el observador en reposo en el mismo marco inercial ( . En la relatividad general, el tiempo transcurre más lentamente en un campo gravitatorio fuerte (
. En la relatividad general, el tiempo transcurre más lentamente en un campo gravitatorio fuerte ( ).
).

Fig. 1.14. Ilustración de la dilatación del tiempo
Dios no sólo expande o contrae, sino que también detiene el tiempo. En el libro de Josué del Antiguo Testamento, está escrito:
“Y el sol se detuvo y la luna se paró, Hasta que la gente se hubo vengado de sus enemigos.” (Josué 10:13)
Este milagro ocurrió durante la batalla de Josué contra los amorreos y demuestra que Dios tiene el poder de congelar el tiempo. Además, Dios realizó un milagro aún más asombroso, según consta en 2 Reyes del Antiguo Testamento:
“Entonces el profeta Isaías clamó a Jehová; e hizo volver la sombra por los grados que había descendido en el reloj de Acaz, diez grados atrás.” (2 Reyes 20:11)
El versículo anterior refleja la respuesta de Dios a la llorosa plegaria del rey Ezequías por una vida más larga. En Su misericordia, Dios escuchó a Ezequías y le concedió 15 años adicionales. Para confirmar Su promesa, Dios realizó una señal milagrosa, haciendo que la sombra de la escalera de Acaz (reloj de sol) retrocediera diez pasos. Este milagro indica que Dios tiene el poder de invertir el tiempo, un concepto que está más allá del alcance de nuestra comprensión científica actual.

Fig. 1.15. Escalera de Ajaz (Reloj de sol)
Para los humanos, el tiempo fluye unidireccionalmente del presente al futuro, pero para Dios, como muestra la Biblia, el tiempo es una variable que Él puede controlar. Dios puede acortar, prolongar, congelar o incluso invertir el tiempo, demostrando así Su soberanía sobre las leyes naturales y poniendo de relieve el contraste entre las limitaciones humanas y Su poder infinito.
e. El Universo bien afinado
El universo finamente sintonizado expresa el hecho de que las constantes físicas fundamentales que conforman y hacen funcionar el universo están finamente torneadas con extrema precisión para que exista vida en el universo
Si la densidad del universo hubiera sido mayor que la densidad crítica, el universo se habría contraído inmediatamente después de su formación. Por el contrario, si hubiera sido menor que la densidad crítica, el universo se habría expandido demasiado rápido, impidiendo la formación de estrellas y galaxias. En cualquiera de los dos casos, no existiríamos en este mundo.
En su libro The Emperor's New Mind, Penrose utilizó la fórmula Bekenstein-Hawking de la entropía de los agujeros negros para estimar las probabilidades en el Big Bang. Calculó que la probabilidad de que el universo llegara a existir de forma que se desarrollara y sustentara la vida tal y como la conocemos es de 1 entre 10 a la potencia de 10123. Esto sugiere que nuestro universo no surgió de un azar o un proceso aleatorio, sino mediante un extraordinario ajuste fino por parte del Creador divino.
Las constantes fundamentales de la física, como la constante gravitatoria, la velocidad de la luz en el vacío, la constante de Planck, la constante de Boltzmann, la constante eléctrica, la carga elemental y la constante de estructura fina, etc., deben estar ajustadas para que exista vida en el universo. Si estas constantes fueran ligeramente diferentes, el universo sería incapaz de albergar vida.
Por ejemplo, si la constante gravitatoria fuera menor de lo que es ahora, la fuerza de la gravedad sería más débil. Esta menor atracción gravitatoria haría imposible que la materia se fusionara en estrellas, galaxias y planetas, incluida la Tierra en la que vivimos hoy. Si la constante de Planck fuera mayor de lo que es ahora, se producirían varios cambios fundamentales en el universo físico. En primer lugar, disminuiría la intensidad de la radiación solar, con lo que llegaría a la Tierra menos energía procedente del Sol. Esta reducción de la energía afectaría a muchos procesos naturales, como el clima y los patrones meteorológicos. Además, valores mayores de la constante de Planck aumentarían el tamaño de los átomos, ya que cambiaría la cuantización de los niveles de energía atómica. Este aumento debilitaría la fuerza de enlace de átomos y moléculas, haciendo que las reacciones químicas fueran menos estables. La fotosíntesis en las plantas, que depende de la absorción precisa de la energía luminosa para convertir el dióxido de carbono y el agua en glucosa, sería menos eficaz. El conjunto de los procesos bioquímicos y físicos que dependen del equilibrio actual de la mecánica cuántica se vería alterado, lo que daría lugar a un entorno radicalmente distinto y menos estable para la vida.
Entre las constantes fundamentales, la constante de estructura fina ha atraído especialmente la atención de los físicos. La constante de estructura fina, denotada por la letra griega cuantifica la fuerza de la interacción electromagnética entre partículas elementales cargadas.
cuantifica la fuerza de la interacción electromagnética entre partículas elementales cargadas.

Es una cantidad adimensional con un valor aproximado de 1/137, una cifra que ha intrigado a los físicos desde su descubrimiento. Su valor exacto es crucial para la estabilidad del universo y la existencia de la vida. Si difiriera mínimamente de su valor actual, la vida tal como la conocemos no existiría.
Si fuera mayor que 1/137, la interacción electromagnética entre las partículas sería más fuerte. Como consecuencia, los electrones estarían más ligados al núcleo, lo que reduciría el tamaño de los átomos y facilitaría la formación de elementos pesados, mientras que los elementos ligeros, como el hidrógeno, tendrían menos probabilidades de formarse. Dado que el hidrógeno es una materia prima crucial para la fusión nuclear, este cambio afectaría directamente a la supervivencia de la vida al limitar la disponibilidad del hidrógeno necesario para la producción de energía en el Sol y las estrellas. Por el contrario, si fuera inferior a 1/137, la interacción electromagnética entre las partículas se debilitaría. Los electrones estarían menos ligados al núcleo, lo que daría lugar a átomos y moléculas inestables. Esta inestabilidad haría que los átomos y las moléculas se descompusieran más fácilmente, impidiendo la formación de moléculas complejas como el ADN y las proteínas, esenciales para la vida. Así pues, cualquier cambio significativo en la constante de estructura fina tendría profundas implicaciones para la formación de la materia y el potencial de vida en el universo.
Desconocemos el origen de su valor numérico ≈ 1/137. Dirac consideraba que el origen de era ‘el problema más fundamental sin resolver de la física’. Feynman describió como un ‘número de Dios’ o ‘número mágico’ que da forma al universo, y que nos llega sin comprenderlo. Se podría decir que la ‘mano de Dios’ escribió ese número, y ‘no sabemos cómo empujó Su lápiz’.
≈ 1/137. Dirac consideraba que el origen de era ‘el problema más fundamental sin resolver de la física’. Feynman describió como un ‘número de Dios’ o ‘número mágico’ que da forma al universo, y que nos llega sin comprenderlo. Se podría decir que la ‘mano de Dios’ escribió ese número, y ‘no sabemos cómo empujó Su lápiz’.
Si reescribimos la ecuación de, puede representar varios cocientes: la velocidad de los electrones con respecto a la velocidad de la luz (es decir, la luz viaja 137 veces más rápido que los electrones), la repulsión electrostática con respecto a la energía de un solo fotón, y el radio clásico del electrón con respecto a la longitud de onda Compton reducida del electrón. Además, la relación entre la fuerza electromagnética y la fuerza gravitatoria es de 10(36), y la relación entre la fuerza electromagnética y la fuerza fuerte es de 1/137. Así pues, el valor numérico de la constante adimensional podría servir como punto de referencia para las cuatro fuerzas fundamentals.
Como se menciona en el capítulo 3, «Física de partículas y creación», toda la materia del universo (baryones) está compuesta por las partículas fundamentales descritas por el Modelo Estándar: quarks, leptones, bosones de gauge y el bosón de Higgs, que suman un total de 17. Cada partícula posee su propia masa, carga y espín únicos. Si alguna de estas propiedades fundamentales fuera ligeramente diferente, las estructuras atómicas, moleculares, biológicas y cósmicas que conocemos no existirían.
Por ejemplo, si se alterara la diferencia de masa entre los quarks up y los quarks down, se rompería el delicado equilibrio que hace que los protones sean estables y los neutrones solo ligeramente más pesados. En tal caso, no se podría formar hidrógeno ni sintetizar núcleos más pesados, lo que haría imposible la existencia de los átomos. Si la masa del electrón fuera significativamente diferente, el tamaño de los átomos y los niveles de energía cambiarían, y ya no se producirían enlaces químicos estables, lo que impediría la formación de moléculas complejas. Si se modificaran las propiedades del bosón de Higgs, se alteraría el mecanismo que da masa a todas las partículas elementales, lo que cambiaría la estructura misma del universo.
Además, si las cargas eléctricas de los protones y los electrones no fueran exactamente iguales y opuestas, los átomos neutros no podrían existir. Si las cargas de los quarks fueran diferentes, las propiedades de los protones y los neutrones cambiarían, lo que socavaría la posibilidad de que existieran los núcleos atómicos. Si los electrones no tuvieran un espín de 1/2, el principio de exclusión de Pauli no se cumpliría y los átomos no podrían mantener su estructura. Del mismo modo, si los bosones no tuvieran valores de espín enteros, se rompería el marco del campo cuántico que permite que operen fuerzas como el electromagnetismo, la fuerza fuerte y la fuerza débil. Por último, si el bosón de Higgs no fuera una partícula de espín 0, el propio mecanismo de generación de masa fallaría y las partículas no podrían existir en su forma actual.
El universo finamente ajustado refleja el asombroso equilibrio y precisión que subyacen a la existencia de todas las cosas. Desde la densidad crítica del universo, establecida con una exactitud inimaginable, hasta el cálculo de Penrose de la probabilidad infinitesimal de tales condiciones iniciales, pasando por los delicados valores de la constante gravitacional, la constante de Planck y la constante de estructura fina, cada detalle apunta a un cosmos exquisitamente calibrado para la vida. Incluso las propias partículas fundamentales —quarks, leptones, bosones y Higgs— poseen precisamente las masas, cargas y espines adecuados para permitir la existencia de átomos, moléculas, estrellas y, en última instancia, seres vivos. Tal armonía no puede atribuirse razonablemente al azar ciego.
Esta extraordinaria precisión no solo inspira asombro, sino que también nos obliga a plantearnos preguntas más profundas sobre el origen y el propósito del universo. La perfecta interacción de las leyes físicas lleva la marca de un diseño intencionado, y el concepto de la creación divina ofrece una explicación profunda y convincente. Al igual que una orquesta solo produce una hermosa sinfonía cuando todos los instrumentos están perfectamente afinados, el universo también da testimonio de la sabiduría y el poder del Creador, que ha ordenado todas las cosas con un propósito y un significado.
Si aquellos que simplemente descubrieron los principios fundamentales del universo —la gravedad, la relatividad, el principio de incertidumbre, el principio de exclusión de Pauli y el mecanismo de Higgs— son honrados como genios y galardonados con premios Nobel, ¿cuánto más grande es Dios, el Creador que no solo concibió estas leyes y principios, sino que también dio origen a todo el universo?
2. La obra maestra de Dios, la Tierra
(¿Qué tan única es la Tierra en el universo?)
La Tierra en la que vivimos ofrece una serie de condiciones muy precisas, esenciales para la supervivencia de los organismos vivos. Estas condiciones son tan precisas que a menudo sirven como extensión del universo ajustado
En este contexto, exploraremos diez condiciones especiales de la Tierra que son particularmente únicas y cruciales para sustentar la vida tal y como la conocemos. Estas condiciones ponen de relieve el extraordinario equilibrio y precisión necesarios para sustentar los organismos vivos, haciendo de nuestro planeta un oasis excepcional en la vasta extensión del universo. Al examinar estos atributos únicos, podemos apreciar mejor la intrincada interacción de los factores que permiten que la vida prospere en la Tierra.
a. Distancia adecuada al Sol
La presencia de agua líquida es crucial para la vida. Para tener agua líquida, un planeta debe orbitar dentro de una región específica alrededor de su estrella central. Si el planeta está demasiado cerca de la estrella, toda el agua hervirá, y si está demasiado lejos, toda el agua se congelará. El rango de órbitas donde el agua ni hierve ni se congela se denomina ‘zona habitable’. La zona habitable estimada en nuestro sistema solar se sitúa entre 0,95 UA y 1,15 UA (1 UA es la distancia de la Tierra al Sol). Por tanto, si la Tierra estuviera un 5% más cerca o un 15% más lejos del Sol, no estaríamos aquí.
El porcentaje de la zona habitable que ocupa el plano de la eclíptica estirado hasta Neptuno (30 UA) es sólo del 0,05%. La excentricidad de la órbita terrestre es otro factor importante que afecta al alcance de la zona habitable. Por ejemplo, si la excentricidad fuera superior a 0,5, toda el agua herviría dos veces al año cerca del perihelio y se congelaría dos veces al año cerca del afelio. Afortunadamente, la excentricidad de la Tierra es sólo de 0,017, lo que da lugar a una órbita casi circular.

Fig. 2.1. Zona habitable (verde) en el sistema solar
b. La inclinación axial adecuada
El eje de rotación de la Tierra está inclinado unos 23,5 grados. Gracias a ello, podemos tener cuatro estaciones y un clima suave. ¿Qué ocurriría si el eje de rotación no estuviera inclinado (0 grados, véase la inclinación axial de Mercurio= 0,0 grados) o estuviera completamente inclinado (90 grados, véase la inclinación axial de Urano = 82,2 grados)?
Si el eje de rotación de la Tierra no estuviera inclinado, se producirían varios cambios significativos en cuanto al clima, las estaciones y la habitabilidad. El ecuador recibiría luz solar directa y constante durante todo el año, lo que daría lugar a temperaturas eternamente cálidas. Por el contrario, los polos recibirían siempre una luz solar mínima, lo que provocaría un frío perpetuo. Este drástico contraste de temperaturas afectaría significativamente a los climas globales y a los patrones meteorológicos.
La ausencia de estaciones tendría profundas repercusiones en los ecosistemas y la agricultura. Las regiones cercanas al ecuador podrían volverse demasiado cálidas para que prosperaran muchos cultivos y organismos, mientras que las regiones polares seguirían siendo inhóspitamente frías. Las latitudes medias se convertirían en las principales zonas habitables, pero incluso estas zonas carecerían de las variaciones estacionales de las que dependen muchas plantas y animales para sus ciclos vitales y su reproducción.
Las sociedades humanas se enfrentarían a graves problemas, como la reducción de la productividad agrícola y el aumento de la presión sobre las tierras habitables. La falta de señales estacionales también podría perturbar las actividades culturales y económicas que dependen del cambio de las estaciones. En general, una Tierra sin inclinación daría lugar a un entorno menos dinámico y menos hospitalario para la vida.

Fig. 2.2. Inclinación axial de la Tierra. Sin inclinación (izquierda) e inclinación de 90 grados (derecha)
Si el eje de rotación de la Tierra se inclinara completamente 90 grados, tendría efectos profundos y dramáticos sobre el clima y el medio ambiente del planeta. En este escenario, un hemisferio experimentaría luz diurna continua durante la mitad del año, mientras que el otro estaría en constante oscuridad, y luego la situación se invertiría durante la otra mitad del año.
Cada hemisferio sufriría variaciones estacionales extremas. Durante el verano, un hemisferio recibiría luz solar constante, lo que provocaría periodos prolongados de calor intenso y condiciones potencialmente desérticas. Por el contrario, durante el invierno, el mismo hemisferio experimentaría una oscuridad continua y temperaturas bajo cero.
Los cambios drásticos de luz y temperatura perturbarían gravemente los ecosistemas. Muchas plantas y animales están adaptados al ciclo estacional actual, y cambios tan extremos amenazarían su supervivencia.
La agricultura, que depende de la previsibilidad de las estaciones, se vería muy afectada. Las regiones actualmente aptas para la agricultura podrían volverse inhabitables, lo que provocaría escasez de alimentos y la necesidad de grandes adaptaciones en las prácticas agrícolas.
En general, un eje completamente inclinado haría la Tierra mucho menos hospitalaria para la vida, creando condiciones ambientales extremas e inestables.
c. El período de rotación y el período orbital adecuados
El periodo de rotación de la Tierra es de 24 horas, con unas 12 horas de día y 12 horas de noche. Nuestro biorritmo está determinado por el periodo de rotación de la Tierra. El periodo de rotación de 24 horas proporciona un bloque de tiempo óptimo para 8 horas de trabajo, 8 horas de sueño y 8 horas de ocio. Sin embargo, no todos los planetas del sistema solar tienen un periodo de rotación óptimo. Por ejemplo, el periodo de rotación de Júpiter es de unas 10 horas, mientras que el de Venus es de 243 días.
Si el periodo de rotación de la Tierra se redujera a 10 horas, ello afectaría significativamente al medio ambiente y a la vida del planeta. Una rotación más rápida acortaría los días y las noches, provocando una rápida alternancia entre luz diurna y oscuridad. Esto podría alterar los ritmos circadianos de muchos organismos, afectando a los patrones de sueño, los comportamientos alimentarios y los ciclos de reproducción.
El aumento de la velocidad de rotación también provocaría efectos Coriolis más fuertes, intensificando los patrones meteorológicos y causando potencialmente tormentas y huracanes más severos. La rotación más rápida también podría afectar a la actividad tectónica de la Tierra. El aumento de la fuerza centrífuga podría provocar terremotos y erupciones volcánicas más frecuentes e intensas.
Por otra parte, si el periodo de rotación de la Tierra fuera de 243 días como en Venus, las consecuencias para el planeta y sus habitantes serían drásticas. Una rotación tan lenta significaría días y noches extremadamente largos, cada uno de unos 120 días.
El lado orientado hacia el Sol experimentaría un calentamiento prolongado, lo que provocaría temperaturas abrasadoras, mientras que el lado opuesto soportaría una oscuridad prolongada y un enfriamiento severo, pudiendo llegar a congelarse. Estas temperaturas extremas dificultarían la supervivencia de la mayoría de las formas de vida. Los prolongados periodos de calentamiento y enfriamiento alterarían la circulación atmosférica, lo que probablemente provocaría fenómenos meteorológicos extremos. Los huracanes, las tormentas masivas y las sequías o inundaciones prolongadas podrían llegar a ser habituales.
Los largos periodos de luz y oscuridad alterarían gravemente los ciclos vitales de plantas y animales, afectando a la fotosíntesis, la reproducción y los patrones de alimentación.
Las actividades humanas, la agricultura y las infraestructuras necesitarían una adaptación significativa para hacer frente a las duras y variables condiciones, lo que supondría un tremendo reto para la supervivencia y la vida cotidiana.
El periodo orbital de la Tierra también es importante para la supervivencia humana. El periodo orbital de la Tierra es de 365 días, con 3 meses para primavera, verano, otoño e invierno. La duración de cada estación está bien equilibrada, asegurando que ninguna estación sea demasiado corta o demasiado larga. Este equilibrio es crucial para los ciclos agrícolas, el crecimiento de las plantas, el calendario de las migraciones animales y otros procesos ecológicos.
¿Qué ocurriría si la Tierra tuviera un período orbital corto, de 88 días, similar al de Mercurio? En este escenario, cada estación duraría sólo unas 3 semanas. La mayoría de los cultivos de la Tierra requieren de 6 a 9 meses desde la siembra en primavera hasta la cosecha en otoño. Sin embargo, con estaciones que cambian cada 3 semanas, los cultivos no tendrían tiempo suficiente para madurar, lo que provocaría una grave escasez de alimentos y afectaría directamente a la supervivencia humana.
Por el contrario, ¿qué ocurre si la Tierra tiene un periodo orbital largo, como 164 años, similar al de Neptuno? Cada estación duraría unos 40 años. Los veranos prolongados provocarían olas de calor y desertización, mientras que los inviernos prolongados causarían largos periodos de frío y hielo, lo que afectaría a la agricultura y los ecosistemas. Mientras que los humanos podrían adaptarse para evitar la escasez de alimentos, los animales salvajes tendrían dificultades para encontrar comida durante un invierno de 40 años. Las duras condiciones prolongadas harían casi imposible la supervivencia de la mayoría de los animales salvajes, provocando una extinción generalizada.
d. La Tierra de tamaño adecuado
Puede que no hayas pensado en ello, pero el tamaño de la Tierra es crucial para la supervivencia de los seres humanos. El tamaño del planeta afecta a su atracción gravitatoria, que a su vez influye en todo, desde la retención de una atmósfera que sustente la vida hasta la capacidad de albergar masas de agua estables y mantener un campo magnético protector.
Si la Tierra tuviera la mitad de su tamaño actual, la gravedad se reduciría a la mitad de la actual. La gravedad reducida tendría impactos significativos y potencialmente devastadores en la capacidad del planeta para albergar vida. La gravedad reducida podría no ser lo suficientemente fuerte como para retener una atmósfera densa. Esta atmósfera más fina ofrecería menos protección contra la radiación solar dañina y los meteoroides y podría no soportar los patrones climáticos estables necesarios para la vida.
La reducción de la gravedad también afectaría a la retención de agua líquida, lo que provocaría un aumento de las tasas de evaporación y, potencialmente, una pérdida de agua superficial con el tiempo. Esto dificultaría el mantenimiento de océanos, ríos y lagos, que son cruciales para sustentar diversos ecosistemas y la civilización humana.
Además, una Tierra más pequeña tendría un campo magnético disminuido, ofreciendo menos protección contra el viento solar. Esto podría despojar a la atmósfera y exponer aún más la superficie a la dañina radiación cósmica y solar, haciendo el planeta mucho menos hospitalario para los seres humanos y otras formas de vida.
Si la Tierra tuviera el doble de su tamaño actual, los efectos sobre la gravedad y la velocidad de escape serían significativos y tendrían profundas implicaciones para la vida en el planeta. La gravedad aumentaría, haciendo que todo en la Tierra se sintiera más pesado, y la velocidad de escape también se duplicaría. Este aumento de la gravedad haría que el movimiento fuera más agotador para los seres humanos y otros organismos, lo que podría provocar un mayor estrés físico y adaptaciones con el paso del tiempo.
La combinación de una mayor gravedad y velocidad de escape también afectaría a la atmósfera. Una mayor atracción gravitatoria retendría más gases, incluidos los tóxicos como el metano y el amoníaco, similares a las atmósferas de Saturno y Júpiter. Estos gases podrían acumularse hasta niveles nocivos, creando un entorno tóxico inadecuado para la mayoría de las formas de vida.
Además, el aumento de la gravedad podría afectar a los procesos geológicos, provocando una actividad volcánica más intensa y montañas más altas. En general, una Tierra más grande, con mayor gravedad y velocidad de escape, plantearía importantes retos para la supervivencia de la vida y podría dar lugar a un entorno más hostil e inestable.

Fig. 2.3. Comparación de los tamaños de los planetas
e. La Tierra con un campo magnético fuerte
La Tierra está rodeada por un sistema de campos magnéticos conocido como magnetosfera, que protege al planeta de la dañina radiación solar y cósmica. Este escudo protector es crucial para mantener la vida en la Tierra. Para tener una magnetosfera, son esenciales dos factores: la velocidad de rotación adecuada y la existencia de un núcleo externo líquido metálico. Afortunadamente, la Tierra posee ambos. La rotación del planeta induce movimientos fluidos (convección) dentro del núcleo externo líquido, generando fuertes campos magnéticos que forman la magnetosfera.
¿Qué pasaría si no tuviéramos magnetosfera? Si la Tierra no tuviera magnetosfera, las consecuencias para los organismos vivos y la atmósfera serían graves. Sin este escudo protector, la dañina radiación solar y cósmica bombardearía el planeta, aumentando significativamente el riesgo de cáncer y mutaciones genéticas en los organismos vivos. Además, la magnetosfera ayuda a evitar la pérdida de atmósfera desviando las partículas cargadas del viento solar. Sin ella, estas partículas despojarían la atmósfera con el tiempo mediante un proceso de pulverización catódica, agotando gases esenciales como el oxígeno y el nitrógeno. Esta erosión atmosférica provocaría un adelgazamiento de la atmósfera, una reducción de la presión superficial y variaciones extremas de temperatura, lo que haría la Tierra menos hospitalaria para la vida.

Fig. 2.4. La magnetosfera de la Tierra desvía los rayos cósmicos
La intensidad del campo magnético de Marte es aproximadamente el 0,01% de la de la Tierra. Debido a la debilidad del campo magnético, la magnetosfera global no pudo formarse en Marte y, en consecuencia, la mayor parte del aire se eliminó mediante el proceso de pulverización catódica.
Las líneas de campo de la magnetosfera convergen en los polos cercanos al Ártico y al Antártico, lo que provoca un debilitamiento natural de la intensidad del campo magnético. Esto puede provocar un aumento de la exposición a la radiación solar en estas zonas. Las partículas cargadas de alta energía ionizan y excitan los átomos de la atmósfera superior y producen auroras boreales y australes.
f. La Tierra con una luna excepcionalmente grande
La Tierra tiene una Luna excepcionalmente grande en comparación con otros planetas. La gran Luna desempeña un papel importante en el sustento de la vida en la Tierra. Aunque su influencia no es inmediatamente evidente, contribuye de dos maneras fundamentales. En primer lugar, la Luna estabiliza el eje de rotación de la Tierra, limitando las grandes variaciones en la inclinación axial. En segundo lugar, genera fuerzas de marea que son esenciales para muchos ecosistemas marinos.
En esta sección se examina en primer lugar el efecto estabilizador de la Luna sobre el eje de rotación de la Tierra. El eje de la Tierra está inclinado aproximadamente 23,5 grados, una configuración que produce variaciones estacionales regulares y favorece unas condiciones climáticas estables. La estabilidad a largo plazo de esta inclinación axial se debe en gran medida a la influencia gravitatoria de la Luna, que orbita alrededor de la Tierra a una distancia relativamente cercana.
Entre los planetas terrestres del sistema solar, la Tierra es el único que posee una luna grande. La Luna tiene un radio de aproximadamente 1740 km. Por el contrario, Mercurio y Venus no tienen satélites naturales. Marte tiene dos pequeñas lunas, Fobos y Deimos, llamadas así por personajes de la mitología griega, pero sus radios son de solo unos 10 km, lo que corresponde aproximadamente al 0,6 % del radio de la Luna.
Como se ha comentado anteriormente, la zona habitable del sistema solar se encuentra a aproximadamente 1 unidad astronómica (UA) del Sol. En comparación, el plano eclíptico, a lo largo del cual orbitan los planetas, se extiende hacia afuera hasta aproximadamente 30 UA, llegando a la órbita de Neptuno. En este contexto, la órbita de la Tierra la sitúa relativamente cerca del Sol. Si bien esta proximidad es necesaria para mantener temperaturas superficiales adecuadas para la vida, también da lugar a una fuerte influencia gravitatoria del Sol sobre la dinámica rotacional de la Tierra.
El efecto desestabilizador de la gravedad solar sobre el eje de rotación de un planeta puede observarse en los planetas vecinos. Marte, que carece de una gran luna estabilizadora, ofrece un ejemplo bien estudiado. Como se ilustra en la figura siguiente, Marte experimenta variaciones sustanciales tanto en su inclinación axial como en su excentricidad orbital debido a las perturbaciones gravitacionales solares. Estas variaciones se producen con un período característico de aproximadamente 150 000 años. Durante los últimos seis millones de años, la inclinación axial de Marte ha variado entre unos 15 y 45 grados, mientras que su excentricidad orbital ha oscilado entre 0,01 y 0,11.
Fig. 2.5. Eje de rotación y cambios de excentricidad en
En Venus, un planeta terrestre similar en tamaño a la Tierra pero sin luna, la influencia gravitacional del Sol produce variaciones grandes y caóticas en el eje de rotación del planeta, que oscilan entre aproximadamente 0 y 180 grados. Esto plantea la pregunta de cómo se comportaría el eje de rotación de la Tierra en ausencia de su Luna. Aunque las estimaciones varían entre los distintos estudios, la mayoría indica que, a escala geológica, la inclinación del eje de la Tierra fluctuaría en un rango de aproximadamente 0 a 70 grados. Variaciones de esta magnitud darían lugar a una inestabilidad climática extrema, lo que provocaría tensiones ambientales graves y persistentes que pondrían en grave peligro la supervivencia a largo plazo de la vida compleja. Afortunadamente, sin embargo, el Creador preparó una luna excepcionalmente grande cerca de la Tierra, lo que evita tales desastres.
A diferencia de otros planetas terrestres, ¿cómo llegó la Tierra a poseer una luna tan masiva como satélite? Se han propuesto varias hipótesis para explicar el origen de la Luna. Entre ellas se encuentran la hipótesis del impacto gigante, que propone que un cuerpo del tamaño de Marte colisionó con la Tierra; la hipótesis de la captura, que sugiere que la Luna fue capturada gravitacionalmente desde otro lugar del sistema solar; la hipótesis de la coformación, que postula que la Tierra y la Luna se formaron juntas a partir del mismo material circumplanetario; y la hipótesis de la fisión, que sugiere que la Luna se formó a partir de material que se separó de la Tierra. Entre ellas, estudios recientes se inclinan claramente por la hipótesis del impacto gigante como la explicación más plausible.
Sin embargo, para que un impacto gigante produzca un sistema satelital como el sistema Tierra-Luna, deben cumplirse una serie de condiciones muy específicas. En primer lugar, la masa del impactador y la geometría de la colisión deben estar dentro de un rango muy estrecho. Se estima que el cuerpo impactante, comúnmente conocido como Theia, tenía una masa aproximada del 10 % de la de la Tierra. Esta masa es suficiente para expulsar una gran cantidad de material del manto a la órbita sin alterar la integridad estructural de la Tierra. Además, la colisión debió de producirse en ángulo oblicuo, ya que un impacto frontal no habría generado el disco de escombros necesario para la formación de la Luna.
En segundo lugar, el material expulsado debió condensarse y fusionarse rápidamente. La Luna debió formarse en un plazo de acreción limitado, antes de que los escombros se dispersaran por las fuertes fuerzas de marea de la Tierra o se reacresaran en el planeta.
Por último, la velocidad de rotación de la Tierra inmediatamente después del impacto debió de ser lo suficientemente alta como para permitir que los escombros se estabilizaran en órbitas estables más allá del límite de Roche. En estas condiciones, el material pudo fusionarse en un único satélite grande en lugar de caer de nuevo a la Tierra. Dado que los parámetros que rigen la masa, el ángulo de impacto, la velocidad y el momento deben alinearse dentro de unos límites muy estrechos, la formación de un satélite masivo como la Luna alrededor de un planeta terrestre se considera un resultado poco común en los sistemas planetarios.
Las mareas oceánicas están causadas principalmente por la fuerza gravitatoria de la Luna. Las mareas proporcionan oxígeno al plancton flotante y lo distribuyen por amplias zonas, donde es consumido por pequeños peces. Las mareas también mezclan el agua dulce rica en nutrientes con el agua salada, suministrando estos nutrientes al plancton y a los peces pequeños. Sin las mareas, el agua dulce rica en nutrientes no se mezclaría con el agua salada, lo que provocaría una proliferación incontrolable de algas. Si las algas contienen toxinas, estas floraciones producirán mareas rojas o floraciones de algas nocivas (FAN), que pueden matar peces, aves marinas, mamíferos e incluso seres humanos. Incluso si las algas no son tóxicas, consumen todo el oxígeno del agua a medida que se descomponen, obstruyendo las branquias de los peces y otros seres marinos. Si no existiera la Luna, el ecosistema marino habría sido destruido hace mucho tiempo. Además, no tendríamos marisco, como langosta, gambas y sushi. Sin embargo, incluso si la Tierra tuviera una Luna más pequeña o más grande que su tamaño actual, o si su ubicación estuviera más lejos o más cerca que su posición actual, podríamos seguir enfrentándonos a problemas similares. Fig. 2.6. Marea roja g. La Tierra con Júpiter como vecino Júpiter es el mayor planeta del sistema solar, 11,2 veces más grande y 318 veces más pesado que la Tierra. La presencia de Júpiter es importante para nuestra supervivencia. La Tierra es bombardeada constantemente por meteoritos (en su mayoría asteroides destrozados y fragmentos de cometas). La frecuencia de caída de meteoritos es de un metro una vez cada hora, de unos metros una vez al día, de unos metros a 10 metros una vez al año, de unos diez metros cada década y de unos diez metros a 100 metros una vez cada siglo. Fig. 2.7. Tamaño y frecuencia de los meteoritos que caen sobre la Tierra Fig. 2.8. Árboles derribados por un meteorito que cayó en Tunguska Cuando los meteoritos de menos de 10 metros entran en la atmósfera, la mayoría se queman debido a la fricción y compresión atmosféricas. Sin embargo, si su tamaño es superior a 10 metros, pueden ocurrir sucesos desastrosos. En 1908, un meteorito de unos 55 metros explotó a una altitud de entre 5 y 10 km en la región de Tunguska y aplastó unos 80 millones de árboles en un área de 2.150 km2. Este acontecimiento de Tunguska es el mayor impacto registrado en la historia de la Tierra. Júpiter es vital porque actúa como una aspiradora cósmica, capturando meteoritos y cometas que, de otro modo, podrían impactar contra la Tierra y provocar sucesos catastróficos como el de Tunguska. Las simulaciones indican que Júpiter es unas 5.000 veces más eficaz en la captura de cometas que Earth. Una demostración notable de ello ocurrió en 1994, cuando Júpiter capturó el cometa fragmentado Shoemaker-Levy 9, que tenía un tamaño estimado de unos 1,8 km. Si este cometa hubiera chocado contra la Tierra, podría haber enviado polvo y escombros a la atmósfera, bloqueando la luz solar. Este bloqueo podría durar lo suficiente como para acabar con toda la vida vegetal, provocando la extinción de personas y animales que dependen de las plantas para sobrevivir. Fig. 2.9. Shoemaker-Levy 9 fragmentado y su impacto en Júpiter. h. La Tierra con tectónica de placas La tectónica de placas es la teoría que describe el movimiento a gran escala de la litosfera terrestre, dividida en varias grandes placas tectónicas por los movimientos convectivos del manto. Esta teoría explica muchos fenómenos geológicos, como el movimiento de los continentes, la formación de montañas, los terremotos y la actividad volcánica. La tectónica de placas desempeña un papel crucial en diversos aspectos de los sistemas terrestres que afectan directa e indirectamente a la supervivencia humana. Uno de los aspectos más importantes de la tectónica de placas es la regulación automática del clima de la Tierra a través del ciclo del carbono. El clima de la Tierra viene determinado principalmente por la radiación solar entrante, el albedo de la superficie terrestre y la composición de la atmósfera. Entre ellos, la radiación solar entrante es casi constante durante mucho tiempo. El albedo es una relación entre la radiación entrante y la radiación reflejada. Una fracción significativa de la radiación reflejada de la superficie de la Tierra será absorbida por las moléculas de dióxido de carbono (CO2) en la atmósfera. La radiación absorbida calienta las moléculas de CO2 y la irradia de nuevo en todas las direcciones, y aproximadamente la mitad de ella vuelve a la Tierra en forma de calor. Esta energía térmica atrapada aumenta la temperatura media global de la superficie, lo que se conoce como efecto invernadero. El ciclo del carbono es el proceso mediante el cual el carbono se intercambia entre la atmósfera, los océanos, el suelo, los minerales, las rocas, las plantas y los animales, lo que resulta crucial para regular el clima de la Tierra. El carbono entra en la atmósfera en forma de CO2 procedente de la respiración, la combustión y las erupciones volcánicas. Las plantas absorben CO2 durante la fotosíntesis, convirtiéndolo en materia orgánica, que es consumida por los animales y devuelta a la atmósfera a través de la respiración y la descomposición. En los océanos, CO2 se disuelve y es utilizado por los organismos marinos para formar conchas de carbonato cálcico (CaCO3). Cuando estos organismos mueren, sus caparazones se acumulan en el fondo oceánico, formando rocas sedimentarias. La meteorización de las rocas terrestres también absorbe CO2, formando carbonatos que son arrastrados a los océanos. Este proceso de meteorización depende de la temperatura. Si hay demasiado CO2 en la atmósfera y aumenta la temperatura por efecto invernadero, el proceso de meteorización aumenta y absorbe más CO2. Si se elimina el CO2 de la atmósfera, la temperatura de la Tierra disminuirá. Si la temperatura de la Tierra disminuye, el proceso de meteorización disminuye y se elimina menos CO2 de la atmósfera. Si esto ocurre, entonces el CO2 acumulado produce más efecto invernadero y aumenta la temperatura. Este proceso se denomina ‘ciclo de meteorización de las rocas por dióxido de carbono’. A lo largo de escalas de tiempo geológicas, la actividad tectónica puede empujar estas rocas ricas en carbono hacia el manto terrestre a través de la subducción. El carbono se libera de nuevo a la atmósfera a través de erupciones volcánicas, completando el ciclo. El ciclo de meteorización de las rocas por dióxido de carbono, que depende de la temperatura, regula automáticamente la temperatura de la Tierra a lo largo de escalas de tiempo geológicas. La figura siguiente muestra cómo ha funcionado este ciclo en los últimos 800.000 años: cuando aumenta la cantidad de dióxido de carbono, aumenta la temperatura de la Tierra, y cuando disminuye el dióxido de carbono, disminuye la temperatura de la Tierra. Fig. 2.11. Correlación entre CO2 y temperatura Sin embargo, el ciclo de meteorización de las rocas por dióxido de carbono no funciona si no hay tectónica de placas. En tal caso, el CO2 acumulado no se reciclará y, por tanto, el efecto invernadero se reduce. Si no hay efecto invernadero, la temperatura de la Tierra disminuirá rápidamente y todas las aguas se congelarán. Si todas las aguas se congelan, la energía solar entrante se reflejará debido al gran albedo y, finalmente, la Tierra entrará en una edad de hielo irreversible. Fig. 2.12. CO2 es reciclado por las placas tectónicas Investigaciones recientes sobre la tectónica de placas sugieren que si la Tierra fuera un 20% más grande o más pequeña de lo que es hoy, si la corteza terrestre contuviera un poco más de metales como el hierro y el níquel, o si la corteza fuera más gruesa, la tectónica de placas no habría funcionado como lo hace ahora. En general, la tectónica de placas es un proceso fundamental que sustenta la vida al mantener la estabilidad geológica y medioambiental de la Tierra. i. La Tierra con un Sol de tamaño adecuado El tamaño de la zona habitable (ZH) de un planeta varía en función del tamaño y el tipo de su estrella central. Para las estrellas pequeñas, como las enanas rojas, la HZ está cerca de la estrella porque ésta emite menos luz y calor. Esto hace que la zona HZ sea más estrecha que la que rodea al Sol. Debido a su proximidad, un planeta en la zona habitable de una enana roja podría quedar atrapado por las mareas, al igual que nuestra Luna lo está con la Tierra. Si eso ocurre, el planeta sería incapaz de generar un campo magnético y formar una magnetosfera debido a su lenta rotación. Sin magnetosfera, la radiación nociva de la estrella podría llegar libremente a la superficie del planeta, dañando las células y el ADN. Además, el lado diurno experimentaría luz diurna constante y calor extremo, mientras que el lado nocturno permanecería en oscuridad perpetua y frío extremo. Para las estrellas grandes, como las gigantes azules o rojas, la ZH está mucho más alejada de la estrella. Sin embargo, los planetas de estas zonas se enfrentan a importantes retos. Las estrellas gigantes evolucionan rápidamente debido a su gran masa, quemando rápidamente todo su hidrógeno, expandiéndose hasta convertirse en supergigantes rojas y sufriendo múltiples etapas de fusión hasta formar un núcleo de hierro. Este núcleo acaba por colapsarse, dando lugar a una explosión de supernova y dejando tras de sí una estrella de neutrones o un agujero negro. La vida útil típica de las estrellas gigantes es de sólo unos pocos millones de años, lo que significa que antes de que la estrella explote en una supernova, los habitantes de un planeta en su HZ tendrían que encontrar otro planeta adecuado al que emigrar para su supervivencia. Además, las estrellas gigantes emiten altos niveles de radiación ultravioleta y de rayos X, que pueden ser perjudiciales para el ADN y las células, haciendo que los entornos superficiales de los planetas dentro de la HZ sean menos hospitalarios para la vida. Además, las estrellas gigantes pueden mostrar una variabilidad significativa en su producción de energía, lo que provoca climas inestables en los planetas en órbita. Esta inestabilidad puede provocar fluctuaciones extremas de temperatura, dificultando la supervivencia de la vida. Fig. 2.13. Cambios en las zonas habitables con el la estrella Las zonas habitables (ZH) alrededor de estrellas similares al Sol ofrecen muchas ventajas. Estas estrellas tienen una producción de energía relativamente estable durante largos periodos, proporcionando luz y calor constantes a los planetas de sus zonas habitables. Esta estabilidad favorece el desarrollo de climas y ecosistemas estables. La zona habitable alrededor de las estrellas similares al Sol se encuentra a una distancia moderada, ni demasiado cerca ni demasiado lejos de la estrella. El espectro luminoso de las estrellas similares al Sol es ideal para la fotosíntesis, lo que permite a las plantas y otros organismos fotosintéticos convertir eficazmente la luz solar en energía, formando la base de una cadena alimentaria sostenible. Además, las estrellas similares al Sol suelen tener niveles más bajos de actividad estelar dañina en comparación con estrellas más pequeñas como las enanas rojas. Un menor número de erupciones y una actividad magnética menos intensa significan que los planetas de la zona habitable están menos expuestos a la radiación potencialmente dañina y a la destrucción de la atmósfera. La fracción de estrellas similares al Sol es sólo un pequeño porcentaje, ya que la mayoría de las estrellas son más pequeñas y ligeras que el Sol. El Sol es una sola estrella, pero entre el 50% y el 60% de las estrellas son binarias o sistemas estelares múltiples. La zona habitable en los sistemas estelares múltiples es mucho más restringida debido a las órbitas complejas, la iluminación variable, las perturbaciones gravitatorias y los niveles potenciales de radiación. Fig. 2.14. Distribución de la masa de las estrellas Fig. 2.15. Órbita circumbinaria (arriba) y órbita circumprimaria o circunsecundaria (abajo) en sistemas binarios. j. La distancia adecuada al centro de la galaxia Al igual que la HZ en nuestro sistema solar, existe una Zona Galáctica Habitable (GHZ) dentro de una galaxia donde las condiciones son más favorables para la vida. Las condiciones requeridas para la GHZ incluyen metalicidad, densidad estelar, niveles de radiación y entornos orbitales. La GHZ debe tener una concentración óptima de elementos pesados (elementos más pesados que el helio) necesarios para la formación de planetas terrestres y moléculas orgánicas. Aunque los elementos metálicos son más abundantes en el centro galáctico, esta zona no puede considerarse favorable para la GHZ debido a su elevada densidad estelar, que provoca frecuentes explosiones de supernovas, estallidos de rayos gamma (GRB) y otros fenómenos de alta energía. Una explosión de rayos gamma que se produjera a menos de 10.000 años luz de la Tierra tendría probablemente efectos devastadores sobre la atmósfera, el clima y la biosfera del planeta. Los efectos inmediatos incluirían un aumento de la radiación ultravioleta debido a la destrucción de aproximadamente el 40% de la capa de ozono, mientras que los efectos a largo plazo podrían implicar cambios climáticos significativos y extinciones masivas. Un acontecimiento de este tipo supondría una grave amenaza para la civilización humana y el mundo natural. La destrucción del 40% de la capa de ozono permitiría que el aumento de la radiación UV dañara el ADN 16 veces más. El fitoplancton, base de la cadena alimentaria marina, es especialmente sensible a la radiación UV. Una mayor exposición a los rayos UV puede inhibir su crecimiento y reproducción, provocando un descenso de las poblaciones de fitoplancton. El fitoplancton desempeña un papel crucial en el ciclo del carbono al absorber CO2 durante la fotosíntesis. Una disminución del fitoplancton reduciría esta absorción de carbono, lo que podría agravar la acumulación de CO2 en la atmósfera y potenciar el efecto invernadero. Fig. 2.16. Fitoplancton Existen indicios de que en el pasado se produjeron extinciones masivas en la Tierra provocadas por GRB cercanos. Por ejemplo, algunos científicos creen que la extinción del Ordovícico-Silúrico, ocurrida hace unos 450 millones de años, se debió a un GRB a 6.000 años luz de la Tierra. Otro problema que se plantea en el centro galáctico son los frecuentes encuentros cercanos con otras estrellas. Estos encuentros cercanos provocan importantes perturbaciones gravitatorias que pueden desestabilizar las órbitas y los ejes de rotación de los planetas dentro de los sistemas planetarios. Estas perturbaciones pueden provocar cruces orbitales, colisiones o eyecciones del sistema. La influencia gravitatoria de las estrellas cercanas también podría perturbar las órbitas de los objetos de la Nube de Oort y el Cinturón de Kuiper, enviando un mayor número de cometas y asteroides al sistema solar interior. Esto aumentaría la probabilidad de impactos contra planetas, incluida la Tierra. Las afueras de la Galaxia tienen una baja densidad estelar y no presentan estos problemas, pero hay una cuestión crucial: la baja tasa de explosión de supernovas. Esto da lugar a un medio interestelar que carece de elementos metálicos suficientes para la formación de planetas terrestres, lo que hace que las afueras de la Galaxia sean desfavorables para la GHZ. La región favorable para la GHZ es aquella en la que hay suficientes elementos pesados para la formación de planetas, menos supernovas y otros sucesos peligrosos para entornos seguros para la vida, y zonas menos concurridas para órbitas planetarias estables. Además, existe una región en la que la velocidad orbital de las estrellas coincide con la velocidad patrón de los brazos espirales de la Galaxia, conocida como radio de corotación. Dentro del radio de corotación, las estrellas y sus sistemas planetarios experimentan menos interacciones gravitatorias perturbadoras con los brazos espirales, lo que aumenta la probabilidad de que se mantengan las condiciones de habitabilidad. Teniendo en cuenta todas estas condiciones, GHZ se encuentra entre 23.000 y 29.000 años-luz del centro de la Galaxia. Casualmente, nuestro sistema solar está a 26.000 años-luz del centro de la Galaxia y se encuentra en el centro de GHZ. Fig. 2.17. Zonas Galácticas Habitables en la Galaxia En este capítulo hemos explorado diez condiciones únicas y extraordinarias que hacen de la Tierra un planeta excepcional. Estas condiciones están tan intrincadamente equilibradas y calibradas con precisión que la probabilidad de que ocurran por azar es astronómicamente baja. La exactitud requerida para la distancia de la Tierra al Sol, su inclinación axial, su período de rotación, su campo magnético, su atmósfera y otros factores críticos crea un entorno que es excepcionalmente capaz de albergar vida. Esta combinación de condiciones favorables que se dan simultáneamente en otros lugares del universo sería altamente improbable, lo que subraya aún más el carácter distintivo de la Tierra. Además, la protección y estabilidad de que goza la Tierra -protegida de fenómenos cósmicos dañinos y manteniendo un delicado equilibrio ecológico- subrayan su singularidad entre otros planetas. En conjunto, estos factores apoyan firmemente la idea de que la Tierra fue diseñada intencionadamente por el Creador divino para servir de hábitat a la vida. Este delicado equilibrio de condiciones no es una mera coincidencia, sino que sugiere un diseño intencionado e inteligente, que hace de la Tierra un entorno extraordinario y excepcionalmente adecuado para sustentar la vida.
Fig. 2.10. Las placas que forman la corteza terrestre













3. ¿Creación o evolución?
¿Cómo apareció la vida en la Tierra? ¿Los organismos vivos surgieron a través de una larga serie de procesos químicos y biológicos aleatorios (evolución), o las pruebas apuntan hacia algún tipo de diseño intencionado (creación)?
En este capítulo exploraremos esa pregunta desde múltiples perspectivas. Comenzaremos con el origen de la vida en sí, y luego exploraremos si la teoría de Darwin debería llamarse «Teoría de la Evolución» o si sería más adecuado llamarla «Teoría de la Adaptación Genética». Después, investigaremos si los seres humanos realmente evolucionaron a partir de antepasados simios, y a continuación presentaremos la teoría del diseño inteligente. Por último, a través del prisma de la física de partículas, la posibilidad de vida extraterrestre, el instinto animal y los patrones y principios matemáticos que se encuentran en toda la naturaleza, evaluaremos la validez de la teoría de la evolución.
a. El origen de la vida
Todo el debate sobre la validez del creacionismo frente a la evolución gira en última instancia en torno a una sola pregunta: ¿cómo apareció el primer organismo vivo en la Tierra? Si la vida surgió a través de procesos puramente naturales y no guiados, entonces la evolución queda fundamentalmente validada. Por el contrario, si la vida nunca hubiera podido surgir sin la intervención de un Creador, entonces la creación sigue siendo la única explicación válida.
Todos los organismos vivos están compuestos por células, y todas las células están construidas químicamente a partir de macromoléculas importantes, como los ácidos nucleicos (ADN y ARN), las proteínas, los lípidos y los carbohidratos. Las proteínas, las moléculas funcionales primarias, son largas cadenas de aminoácidos cuya secuencia está determinada por la información genética almacenada en el ADN. Además, los lípidos y los carbohidratos son sintetizados por enzimas que siguen las instrucciones codificadas en el ADN, y estas enzimas son en sí mismas proteínas.
Por lo tanto, incluso para construir la célula más simple, las proteínas son absolutamente esenciales; sin embargo, para producir proteínas, el ADN y el ARN ya deben existir. Sin embargo, en la Tierra primitiva no existía ninguno de estos elementos: ni el ADN, ni el ARN, ni las proteínas, ni las enzimas. El origen natural espontáneo de la vida es, desde el principio, imposible debido a esta dependencia circular.
No obstante, los evolucionistas suelen insistir en que, en la Tierra primitiva, el ARN se formó de alguna manera por sí solo. Este escenario, conocido como la «hipótesis del mundo del ARN», postula que el ARN funcionaba simultáneamente como: una molécula genética (como el ARNm) para almacenar información y una enzima catalítica (ribozima, como el ARNr) para sintetizar proteínas.
Sin embargo, en realidad, una sola molécula de ARN funcional requiere una secuencia de nucleótidos extraordinariamente compleja y precisa, lo que hace que su formación natural aleatoria sea prácticamente imposible (probabilidad prácticamente nula). Incluso si, por una casualidad extraordinariamente improbable, apareciera una molécula de ARN de este tipo, sería tan inestable químicamente que se desintegraría rápidamente en el agua. Además, las capacidades catalíticas de las ribozimas conocidas son extremadamente limitadas y muy inferiores a las de las verdaderas enzimas proteicas. Por encima de todo, sigue existiendo la contradicción fundamental: sin proteínas, no puede haber una replicación eficiente del ARN, y sin ARN, no puede haber síntesis de proteínas. Por lo tanto, la hipótesis del mundo del ARN está plagada de contradicciones internas.
Además, incluso si asumimos que el ARN, las proteínas y el ADN surgieron de alguna manera por casualidad, los pasos posteriores —el autoensamblaje en una célula procariota viva, la evolución hacia células eucariotas, la localización de orgánulos, la diferenciación celular, la formación de tejidos y órganos y, finalmente, la aparición de organismos multicelulares— representan cada uno una secuencia adicional de eventos astronómicamente improbables. En consecuencia, en este capítulo se examinará rigurosamente si la vida podría haber surgido realmente solo por procesos aleatorios.
En primer lugar, revisaremos el clásico experimento de Miller-Urey, al que se suele hacer referencia en los debates sobre los orígenes químicos de la vida. Evaluaremos tanto sus conclusiones como sus limitaciones reconocidas, ya que el experimento solo intentó demostrar que se podían formar compuestos orgánicos simples a partir de precursores inorgánicos en las condiciones que se supone que existían en la Tierra primitiva. A continuación, investigaremos, paso a paso, los siguientes temas: i) formación de aminoácidos, ii) formación de ARN, iii) formación de proteínas, iv) formación de ADN, v) formación de células procariotas, vi) formación de células eucariotas, vii) localización de orgánulos, viii) diferenciación celular, ix) formación de tejidos y órganos, x) formación de organismos multicelulares. A través de este análisis sistemático, determinaremos si todas las etapas que afirma la teoría evolutiva, desde la no vida hasta la vida, podrían haber ocurrido realmente a través de procesos aleatorios y no guiados.
i. Experimento de Miller-Urey
El experimento de Miller-Urey es uno de los estudios más citados en los debates sobre el origen químico de la vida. Su objetivo era comprobar si las moléculas orgánicas simples —en particular los aminoácidos, los componentes básicos de las proteínas— podían surgir de forma natural a partir de sustancias inorgánicas en condiciones similares a las del entorno de la Tierra primitiva. Para llevar a cabo este experimento, Miller y Urey asumieron que la atmósfera primigenia estaba compuesta por metano, amoníaco, hidrógeno y vapor de agua. Hicieron circular esta mezcla de gases a través de un aparato cerrado equipado con una unidad de calentamiento, una bomba de vacío, un dispositivo de descarga eléctrica para simular los rayos y un sistema de enfriamiento. Después de hacer funcionar el sistema durante varios días, informaron de la detección de cuatro aminoácidos entre los productos de la reacción.
Aunque el experimento demostró que se pueden formar algunos aminoácidos a partir de gases simples en condiciones controladas de laboratorio, su relevancia para las condiciones atmosféricas reales de la Tierra primitiva y la idoneidad de su aparato experimental siguen siendo objeto de debate. Por estas razones, el experimento de Miller-Urey merece un cuidadoso reexamen.

Fig. 3.1. Diagrama del experimento de Miller-Urey
El experimento de Miller-Urey utilizó un dispositivo de descarga eléctrica para imitar el rayo natural, pero su dispositivo y el rayo natural difieren significativamente en muchos aspectos. Su dispositivo utiliza un voltaje de 50.000 voltios y genera 250 grados de calor, mientras que el voltaje de un rayo es de 100 millones de voltios y genera 50.000 grados de calor. Las descargas eléctricas del experimento de Miller-Urey eran relativamente continuas y podían mantenerse durante largos periodos, lo que garantizaba un aporte de energía constante para las reacciones químicas. En cambio, los relámpagos no se producen de forma continua, sino esporádica, y su duración es extremadamente breve, de unos pocos microsegundos o milisegundos.
Los cometas son restos del sistema solar primitivo y contienen material de construcción primordial que ha permanecido relativamente inalterado. La composición de los cometas puede proporcionar información valiosa sobre la composición de la atmósfera de la Tierra primitiva. La composición principal de los cometas es agua (86%), dióxido de carbono (10%) y monóxido de carbono (2,6%). El amoníaco y el metano ocupan menos del 1% cada uno. Este resultado sugiere que el gas utilizado en el experimento de Miller-Urey no representa con exactitud la atmósfera de la Tierra primitiva, ya que no contiene el gas más abundante, el dióxido de carbono, ni el segundo más abundante, el monóxido de carbono. Además, el dióxido de carbono de es un agente oxidante que inhibe la formación de aminoácidos.
Composición | Ratio (%) | Referencia |
agua | 100 (86%) | Pinto et al. (2022) |
dióxido de carbono | 12 (10%) | Pinto et al. (2022) |
monóxido de carbono | 3 (2.6%) | Pinto et al. (2022) |
amoníaco | 0.8 (0.7%) | Russo et al. (2016) |
metano | 0.7 (0.6%) | Mumma et al. (1996) |
Tabla 3.1. Composición de los cometas (agua=100)
El experimento de Miller-Urey suponía que la atmósfera prebiótica de la Tierra primitiva era una atmósfera reductora. Sin embargo, si fuera una atmósfera oxidante, dificultaría la formación de aminoácidos al descomponer u oxidar las moléculas orgánicas. Las condiciones de la atmósfera de la Tierra primitiva son objeto de continuo debate e investigación científica. Urey (1952), Miller (1953) y Chyba y Sagan (1997) defienden una atmósfera reductora, mientras que Albeson (1966), Pinto et al. (1980), Zahnle (1986) y Trail et al. (2011) defienden una atmósfera oxidante.
Cabe destacar el trabajo de Trail et al. (2011) publicado en Nature. Analizaron el estado de oxidación de los cristales de circón de la era Hadeica utilizando la relación de los estados de oxidación del cerio (Ce). El análisis indicó que los magmas del Hadeico estaban más oxidados de lo que se pensaba, con condiciones como las de los gases volcánicos modernos. El estado más oxidado de los magmas del Hadeico implica que la desgasificación volcánica habría liberado menos hidrógeno (H2) y más vapor de agua (H2O), dióxido de carbono (CO2) y dióxido de azufre (SO2). Llegaron a la conclusión de que la atmósfera de la Tierra primitiva era probablemente menos reductora y más oxidante de lo que se pensaba tradicionalmente. Sus resultados cuestionaron la validez del experimento de Miller-Urey y sugirieron que quizá no fuera posible formar aminoácidos por abiogénesis en la Tierra prebiótica primitive.
Los aminoácidos producidos en el experimento se recogieron y conservaron en condiciones de laboratorio. En las duras y variables condiciones de la Tierra primitiva, estos compuestos podrían haber sido menos estables y más propensos a la degradación. La concentración de moléculas orgánicas en el experimento se controló y se mantuvo a niveles relativamente altos. En la Tierra primitiva, estas moléculas podrían haber estado muy diluidas en vastos océanos o sometidas a una rápida dispersión, reduciendo potencialmente las posibilidades de una mayor evolución química.
Otro problema clave es la quiralidad. Los aminoácidos producidos eran racémicos, es decir, contenían cantidades iguales de isómeros zurdos y diestros. La vida en la Tierra utiliza principalmente aminoácidos zurdos (99,3%), y el origen de esta homociralidad sigue sin explicarse por el experimento de Miller-Urey. Además, ningún experimento realizado en condiciones simuladas de la Tierra primitiva ha producido nunca los 20 aminoácidos de tipo L simultáneamente. Incluso si los 20 aminoácidos hubieran surgido por casualidad, la probabilidad de que se polimerizaran en las secuencias exactas necesarias para formar proteínas funcionales —o se ensamblaran espontáneamente en ácidos nucleicos como el ARN y el ADN— es estadísticamente insignificante.
Teniendo esto en cuenta, pasamos ahora a una pregunta fundamental: ¿podría el ARN, considerado un componente clave en el origen de la vida, haberse formado de forma natural en el entorno primitivo de la Tierra, o es tal escenario efectivamente imposible?
ii. La formación del ARN
Para comprender lo extraordinariamente difícil que es que el ARN se forme de forma natural, primero debemos entender de qué está compuesto realmente el ARN. El ARN es un polímero largo formado por cientos o miles de nucleótidos unidos entre sí por enlaces fosfodiéster. Cada nucleótido, a su vez, está compuesto por tres componentes distintos: una base nitrogenada (adenina (A), guanina (G), citosina (C) o uracilo (U)), un azúcar de cinco carbonos llamado ribosa y uno o más grupos fosfato.
Para que una sola molécula de ARN funcional surgiera espontáneamente en la Tierra primitiva, primero tendrían que producirse estos tres componentes en cantidad y pureza suficientes en condiciones prebióticas. Luego, en las proporciones correctas y sin reacciones secundarias destructivas, tendrían que ensamblarse en nucleótidos y posteriormente unirse entre sí —cientos o miles de veces— en la secuencia precisa mediante repetidas reacciones de deshidratación-condensación en un entorno acuoso donde tales reacciones son muy poco favorables.
Examinemos ahora, paso a paso, si estos tres componentes esenciales podrían haberse formado de manera plausible en la Tierra primitiva y, de ser así, qué tan probables fueron realmente tales eventos.
- Formación de bases nitrogenadas
Las bases nitrogenadas son moléculas complejas con intrincadas estructuras en anillo. El ensamblaje espontáneo de estas moléculas a partir de compuestos prebióticos más simples es altamente improbable, ya que requiere reacciones químicas específicas, condiciones de reacción concretas y catalizadores para formar las estructuras anulares. Entre ellas se encuentran las reacciones de aminación, en las que se añade un grupo amino (NH2) a una columna vertebral de carbono, que requieren compuestos nitrogenados como el amoníaco y aldehídos o cetonas, a menudo facilitadas por catalizadores o altas temperaturas. Las reacciones de desoxigenación, que eliminan átomos de oxígeno, necesitan agentes reductores como hidrógeno o gases de metano. La formación de anillos, crucial para crear la estructura de base nitrogenada, suele producirse en procesos de varios pasos en condiciones de alta temperatura y alta presión, a menudo catalizados por iones metálicos. Por último, la adición de bases nitrogenadas puede requerir entornos de alta energía y compuestos precursores específicos para completar el proceso.
Se cree que el entorno de la Tierra primitiva variaba enormemente en cuanto a temperatura, pH y compuestos químicos disponibles. Crear las condiciones precisas necesarias para la síntesis de bases nitrogenadas habría sido extremadamente difícil. Por ejemplo, las condiciones de alta energía necesarias para formar estas bases podrían no haber estado presentes o no haberse mantenido de forma constante. Incluso en condiciones de laboratorio optimizadas, el rendimiento de las bases nitrogenadas suele ser bajo. Esto plantea la cuestión de si podrían haberse producido de forma natural cantidades suficientes de estas bases para apoyar la formación de ARN u otros ácidos nucleicos. Las vías que conducen a la síntesis de bases nitrogenadas implican múltiples pasos y compuestos intermedios. La probabilidad de que todas las condiciones y compuestos necesarios estén presentes simultáneamente y en las proporciones correctas es cuestionable.
La formación de bases nitrogenadas suele requerir catalizadores que impulsen las reacciones químicas. En un mundo prebiótico, la presencia de tales catalizadores en las concentraciones y condiciones adecuadas es incierta. Sin estos catalizadores, las velocidades de reacción serían demasiado lentas para ser significativas. Incluso si las bases nitrogenadas pudieran formarse espontáneamente, su estabilidad en un entorno prebiótico es cuestionable. Estas moléculas son propensas a degradarse bajo la radiación UV, la hidrólisis y otros factores ambientales. Esta inestabilidad dificultaría su acumulación y posterior uso en la formación de ARN.
- Formación del azúcar ribosa
La reacción de la formosa, que implica la polimerización del formaldehído en presencia de un catalizador, puede producir ribosa. Esta reacción carece de especificidad, lo que conduce a un bajo rendimiento de ribosa en relación con otros azúcares. También requiere condiciones específicas, como la presencia de hidróxido de calcio como catalizador, que puede no haber estado universalmente disponible o no haber sido estable en entornos prebióticos. Para que la ribosa fuera útil en la síntesis prebiótica de ARN, tendría que sintetizarse y estabilizarse selectivamente. Sin embargo, la reacción de la formosa no favorece la formación selectiva de ribosa, y la mezcla de azúcares resultante complica la utilización de ribosa para la síntesis de ARN. Habría sido necesaria la presencia de mecanismos para estabilizar la ribosa o seleccionarla a partir de una mezcla compleja. Se han propuesto posibles agentes estabilizadores, como los minerales de borato, pero su disponibilidad y eficacia en condiciones prebióticas son inciertas.
La reacción de la formosa requiere formaldehído, que debe estar presente en concentración suficiente. La producción y estabilidad del formaldehído en condiciones prebióticas no son posibles, ya que el formaldehído puede polimerizarse fácilmente o reaccionar con otros compuestos. Es posible que las condiciones ambientales específicas necesarias para que la reacción de la formosa se produzca de forma eficiente y produzca ribosa (por ejemplo, pH óptimo, temperatura, presencia de catalizadores) no fueran frecuentes ni estables en la Tierra primitiva. Incluso en condiciones controladas de laboratorio, el rendimiento de la ribosa es bajo y la reacción produce una mezcla compleja de azúcares, lo que pone de manifiesto el reto que supone aislar la ribosa en un entorno prebiótico.
La ribosa es un azúcar pentosa químicamente inestable y propenso a una rápida degradación, sobre todo en las condiciones que se cree prevalecían en la Tierra primitiva. La inestabilidad se debe a que la ribosa se hidroliza fácilmente en soluciones acuosas y puede degradarse mediante procesos como la reacción de Maillard y la caramelización. Además, los estudios han demostrado que la ribosa tiene una vida media corta, especialmente en condiciones alcalinas, lo que hace improbable que se acumule en cantidades significativas a lo largo de escalas de tiempo geológicas.
- Formación del grupo fosfato
La formación de grupos fosfato en condiciones prebióticas se enfrenta a desafíos porque las fuentes de fosfato fácilmente disponibles eran relativamente escasas en la Tierra primitiva. El fosfato suele encontrarse en minerales como la apatita, que no son muy solubles en agua, lo que dificulta la libre disponibilidad del fosfato en los entornos acuosos en los que se cree que tuvo lugar la química prebiótica. Los minerales de fosfato tienden a ser químicamente inertes en condiciones de pH neutro. Esta baja reactividad supone una barrera importante para la incorporación del fosfato a las moléculas orgánicas necesarias para la vida.
La formación de ésteres de fosfato, críticos para la síntesis de nucleótidos, requiere un aporte energético significativo. En condiciones prebióticas, las fuentes de energía y los procesos catalíticos necesarios para superar estas barreras habrían sido limitados. Algunos estudios han demostrado que las condiciones de alta energía, como las creadas por la caída de rayos o la actividad volcánica, pueden facilitar la formación de moléculas que contienen fosfatos. Sin embargo, estos escenarios requieren condiciones específicas y transitorias que pueden no haber estado muy extendidas.
La formación de polifosfatos, que son cadenas de grupos fosfato, suele requerir condiciones específicas, como altas temperaturas o la presencia de catalizadores que podrían no haber estado fácilmente disponibles en entornos prebióticos. Los polifosfatos son propensos a la hidrólisis, descomponiéndose en compuestos de fosfato más simples. La estabilidad de estos compuestos en las condiciones fluctuantes de la Tierra primitiva es cuestionable.
Aunque algunos experimentos han demostrado la formación de moléculas que contienen fosfatos en condiciones prebióticas simuladas, a menudo requieren condiciones muy específicas y controladas que pueden no reflejar de forma realista los entornos de la Tierra primitiva. Además, los rendimientos de las moléculas que contienen fosfatos en los experimentos de síntesis prebiótica suelen ser bajos, lo que plantea dudas sobre la eficacia y plausibilidad de que estos procesos se produzcan en una Tierra prebiótica a escalas suficientes para impulsar el origen de la vida.
- Formación de nucleótidos de ARN funcionales
Aunque se hayan superado todos los retos y se hayan creado con éxito las bases nitrogenadas, el azúcar ribosa y los grupos fosfato, queda otro obstáculo importante: la formación de nucleótidos de ARN funcionales.
Existen muchos tipos de ARN: ARN implicados en la síntesis de proteínas (ARNm, ARNr, ARNt, etc.), ARN implicados en la modificación postranscripcional (ARNsn, ARNsno, etc.), ARN reguladores (ARNa, ARNmi, etc.) y ARN parásitos. El número de nucleótidos de las moléculas de ARN depende de su tipo. Algunos ejemplos son
- ARNm y ARNr - de cientos a miles
- ARNt - 70 a 90
- snRNA - 100 a 300
- miARN - 20 a 25.
Supongamos que la molécula típica de ARN, para la que queremos estimar la probabilidad de formación, tiene 100 nucleótidos de longitud. En ese caso, cada posición de la secuencia de ARN puede estar ocupada por una de cuatro bases: adenina, uracilo, citosina o guanina. El número total de secuencias posibles de 100 nucleótidos de longitud es 4100 (=1,6x1060) y la probabilidad de formar ARN funcional es 1/1,6x1060 = 6,2x10-61. Esta probabilidad extremadamente pequeña sugiere que el ARN funcional no puede formarse espontáneamente, incluso en presencia de bases nitrogenadas preexistentes, azúcar ribosa y grupos fosfato.
iii. La formación de proteínas
La formación de proteínas implica la síntesis de aminoácidos, su polimerización en péptidos y el plegamiento de estos péptidos en proteínas funcionales. Examinemos los problemas y retos de estos procesos en condiciones prebióticas.
Las proteínas están compuestas por largas cadenas de aminoácidos, denominadas cadenas polipeptídicas, dispuestas en secuencias muy específicas. El número de aminoácidos de una sola proteína puede oscilar entre varias docenas y varios miles. Por ejemplo, la pequeña proteína insulina contiene unos 51 aminoácidos, la proteína mediana mioglobina unos 153 aminoácidos, la gran proteína hemoglobina unos 574 aminoácidos y la proteína gigante titina unos 34.350 aminoácidos. Es casi imposible formar largas cadenas peptídicas mediante un proceso aleatorio a partir de una combinación de 20 tipos de aminoácidos. Por ejemplo, la probabilidad de formar una cadena polipeptídica en la pequeña proteína insulina a través de un proceso aleatorio es 1/2051 = 4.4x10-67 ≈ 0.
Aunque las cadenas polipeptídicas se formaran de algún modo, deben plegarse en estructuras tridimensionales específicas para ser proteínas funcionales. El proceso de plegamiento de una cadena polipeptídica en una proteína funcional implica varios pasos clave, cada uno de ellos impulsado por diversas interacciones químicas y asistido por maquinaria molecular dentro de la célula.
Las secciones de la cadena polipeptídica (estructura primaria) se pliegan en estructuras secundarias conocidas como hélices alfa y láminas beta. Estas estructuras se estabilizan mediante enlaces de hidrógeno entre los átomos del esqueleto de la cadena polipeptídica. Otras estructuras secundarias, como giros y bucles, conectan las hélices y las láminas, contribuyendo al plegamiento global de la proteína. Las estructuras secundarias se pliegan aún más en una forma tridimensional específica, conocida como estructura terciaria. Este proceso es impulsado por interacciones hidrofóbicas, en las que las cadenas laterales no polares se agrupan lejos del medio acuoso, impulsando al polipéptido a plegarse en una forma compacta y globular; enlaces de hidrógeno, que se forman entre las cadenas laterales polares y la columna vertebral, estabilizando la estructura plegada; enlaces iónicos, con interacciones electrostáticas entre cadenas laterales de carga opuesta que contribuyen a la estabilidad de la proteína ; y enlaces disulfuro, en los que enlaces covalentes entre residuos de cisteína proporcionan estabilidad adicional a la estructura.
En algunas proteínas con múltiples cadenas polipeptídicas (subunidades), estas unidades plegadas se unen para formar la estructura cuaternaria. Para evitar errores, las proteínas chaperonas ayudan en el proceso de plegamiento impidiendo el plegamiento incorrecto y la agregación. Ayudan a la cadena polipeptídica a alcanzar su conformación correcta. La proteína puede sufrir cambios y correcciones conformacionales menores para alcanzar su conformación más estable y funcional. Pueden producirse modificaciones químicas, como la fosforilación, la glicosilación o la escisión, que estabilizan aún más la proteína o la preparan para su función específica.
La formación de enlaces peptídicos entre aminoácidos requiere una importante cantidad de energía. En condiciones prebióticas, la disponibilidad de fuentes de energía constantes y suficientes para impulsar estas reacciones es cuestionable. Aunque se han propuesto varias fuentes de energía, como los rayos, la radiación ultravioleta y el calor volcánico, la eficacia y fiabilidad de estas fuentes para facilitar la formación de enlaces peptídicos son discutibles. Es probable que las condiciones de la Tierra primitiva fueran duras y variables, con temperaturas extremas, niveles de pH y cambios ambientales. Estas condiciones podrían haber perturbado el delicado proceso de formación de enlaces peptídicos y la estabilidad de los péptidos formados.
Los péptidos y aminoácidos están sujetos a hidrólisis y degradación en medios acuosos. La estabilidad de los péptidos formados durante largos periodos es motivo de preocupación, ya que podrían degradarse más rápido de lo que se forman. La falta de mecanismos de protección en condiciones prebióticas significa que los péptidos recién formados podrían descomponerse rápidamente por factores ambientales como la radiación UV y las fluctuaciones térmicas. Aunque las superficies minerales como las arcillas pueden catalizar la formación de enlaces peptídicos, la eficacia, especificidad y rendimiento de estas reacciones en condiciones naturales no están bien demostradas. No se sabe con certeza hasta qué punto estas superficies pueden producir una amplia gama de péptidos necesarios para la vida. Las condiciones precisas en las que se producen estas reacciones catalizadas por minerales (temperatura, pH, etc.) deben estar estrechamente controladas, y es posible que tales condiciones no se dieran en la Tierra primitiva. Algunos experimentos que demuestran la formación de péptidos se realizaron en condiciones muy controladas, pero es posible que no reflejen con exactitud las condiciones caóticas y variables de la Tierra primitiva.
La hipótesis del mundo del ARN postula que las moléculas de ARN catalizaron la formación de péptidos. Sin embargo, la aparición simultánea de ARN y péptidos funcionales plantea el problema del ‘huevo y la gallina’, ya que ambos son interdependientes.Sin ARN, no pueden formarse proteínas.
Las proteínas requieren aminoácidos con la misma quiralidad (L-aminoácidos). La síntesis prebiótica suele producir mezclas racémicas que contienen cantidades iguales de isómeros izquierdos y derechos. La formación espontánea de proteínas homocirales a partir de tales mezclas es estadísticamente improbable.

Fig. 3.2. Síntesis de proteínas
iv. La formación del ADN
La formación del ADN en condiciones prebióticas es un proceso complejo y especulativo que implica varios pasos clave, como la síntesis de nucleótidos, la formación de cadenas de polinucleótidos, el emparejamiento de bases, la formación de la doble hélice, la condensación del ADN y la replicación y asistencia enzimática.
Al igual que el ARN, los nucleótidos del ADN se componen de tres partes: bases nitrogenadas (adenina, guanina, citosina, timina), azúcar desoxirribosa y grupos fosfato. El nivel de dificultad para la formación espontánea del ADN será comparable al del ARN. Una dificultad adicional para el ADN es la formación de su estructura de doble hélice. La estructura de doble hélice del ADN depende del emparejamiento preciso de bases entre la adenina y la timina, y entre la citosina y la guanina. Lograr esta especificidad de forma espontánea, sin una plantilla o mecanismo que la guíe, es extremadamente improbable. Para obtener una doble hélice estable, los nucleótidos deben estar dispuestos en un orden específico, con secuencias complementarias en las hebras opuestas. La probabilidad de formar espontáneamente dos secuencias complementarias que se alineen perfectamente es extremadamente baja.
La replicación del ADN requiere complejas enzimas y maquinaria proteica para garantizar su precisión y fidelidad. La lista de enzimas clave que intervienen en la replicación del ADN incluye la helicasa, las proteínas de unión de una sola hebra (SSB), la primasa, la ADN polimerasa, la ribonucleasa H (RNasa H), la ADN ligasa y la topoisomerasa. La formación espontánea de una doble hélice no incluiría estos componentes esenciales, por lo que la replicación y la corrección de errores serían altamente improbables. Sin mecanismos de corrección de errores, cualquier ADN formado espontáneamente probablemente acumularía errores rápidamente, comprometiendo su estabilidad y funcionalidad.
El número total de aminoácidos en las enzimas típicas que participan en la replicación del ADN oscila entre cientos y varios miles. La probabilidad de producir cualquiera de estas enzimas por azar es prácticamente nula. Por ejemplo, la probabilidad de producir RNasa H por azar es de sólo 20-155 o 2,2x10-202 ≈ 0. Esta probabilidad increíblemente pequeña está esencialmente fuera del ámbito de la ocurrencia práctica y nunca ocurrirá en la naturaleza.
Incluso si el ADN se formara de alguna manera, tendría que pasar por un proceso de condensación del ADN muy complejo. El proceso de condensación del ADN transforma una molécula de ADN larga y lineal en una estructura muy compacta y organizada capaz de encajar en el núcleo celular. El proceso de condensación es esencial para el almacenamiento, la protección y la regulación eficaces del ADN, así como para la correcta segregación de los cromosomas durante la división celular. Este proceso implica la formación de nucleosomas, fibra de 30 nm, dominios en bucle, plegamiento de orden superior y cromosomas metafásicos.
El nucleosoma puede formarse si el ADN se enrolla alrededor de las proteínas histonas. Cada nucleosoma está formado por unos 147 pares de bases de ADN enrollados alrededor de un octámero de histonas (dos copias de cada una de las proteínas H2A, H2B, H3 y H4). La estructura resultante de se parece a las cuentas de un collar, con nucleosomas (las cuentas) conectados por ADN enlazador (el collar).
La cadena del nucleosoma se enrolla aún más en una fibra más compacta de 30 nm, facilitada por la histona H1 enlazadora, que se une al nucleosoma y al ADN enlazador. La fibra de 30 nm puede adoptar una configuración en solenoide o en zigzag, dependiendo de las interacciones del nucleosome.
La fibra de 30 nm forma dominios en bucle uniéndose a un andamiaje proteico dentro del núcleo. El andamiaje o las regiones de unión a la matriz (SARs/MARs) anclan estos bucles. Estos bucles, que suelen tener una longitud de 40-90 pares de kilobases (kb), proporcionan una mayor compactación y desempeñan un papel en la regulación génica al acercar elementos reguladores distantes a los genes.
Los dominios en bucle se pliegan aún más formando fibras más gruesas, conocidas como fibras de cromonema. Estas fibras se enrollan y pliegan aún más, lo que da lugar a una estructura más condensada.
Durante la división celular, especialmente en metafase, la cromatina alcanza su máximo nivel de condensación para formar cromosomas visibles. Esto implica la acción de proteínas condensinas que ayudan a superenrollar y compactar la cromatina. Cada cromosoma está formado por dos cromátidas hermanas idénticas que se mantienen unidas en el centrómero, lo que garantiza una segregación precisa durante la división celular.
El grado de condensación influye en la expresión génica: la heterocromatina muy condensada es inactiva desde el punto de vista transcripcional, mientras que la eucromatina poco condensada es activa. Una condensación adecuada es crucial para la segregación precisa de los cromosomas durante la mitosis y la meiosis.
Como se ha visto anteriormente, la formación y replicación del ADN son altamente complejas, requieren una coordinación bioquímica precisa y la participación de varias enzimas. Sin embargo, la teoría evolutiva no ofrece una explicación clara de cómo se originaron estos mecanismos, limitándose a afirmar que el ADN evolucionó a partir del ARN sin abordar los retos críticos. Para que esta afirmación sea válida, debe explicar cómo se formó el ARN, cómo surgió la estructura de doble hélice del ADN y cómo se originaron las enzimas de replicación esenciales. Sin estas respuestas, la idea sigue siendo especulativa. Teniendo en cuenta estos factores, la formación del ADN es el resultado de un diseño intencionado y no del azar.

Fig. 3.3. Proceso de replicación del ADN
v. La formación de las células procariotas
Para continuar nuestra discusión, supongamos que el ARN, las proteínas y el ARN se produjeron espontáneamente. Entonces, el siguiente paso hacia la vida es la formación de células. Existen dos tipos principales de células: procariotas y eucariotas. Las células procariotas, que se encuentran en organismos como las bacterias y las arqueas, son más simples y carecen de un núcleo definido. Su material genético está contenido en una única molécula circular de ADN que flota libremente en el citoplasma. Las células procariotas también carecen de orgánulos unidos a membranas. Las células eucariotas, presentes en plantas, animales, hongos y protistas, tienen una estructura más compleja. Contienen un núcleo definido rodeado por una membrana nuclear. Las células eucariotas también poseen varios orgánulos membranosos, como las mitocondrias, el retículo endoplásmico y el aparato de Golgi, que desempeñan funciones específicas esenciales para la supervivencia y el buen funcionamiento de la célula.
Los científicos afirman que las protocélulas evolucionaron hasta convertirse en células procariotas mediante un proceso gradual impulsado por la selección natural, la mutación y la adaptación al entorno. La existencia de protocélulas, hipotéticas precursoras de las células modernas, se enfrenta a varias críticas importantes. Una de las principales es la formación espontánea de bicapas lipídicas, esenciales para crear un entorno cerrado y estable. Las condiciones necesarias para formar y mantener estas bicapas de forma constante en la Tierra primitiva son altamente especulativas. Además, la integración de componentes funcionales, como ARN o proteínas simples, dentro de estas estructuras lipídicas requiere interacciones muy específicas que son estadísticamente improbables sin algún mecanismo de guía. Además, la capacidad de las protocélulas para replicarse y evolucionar, una característica clave de los organismos vivos, carece de suficiente apoyo experimental, lo que plantea interrogantes sobre su papel en el origen de la vida. Por estas razones, las primeras células que aparecieron en la Tierra habrían sido células procariotas.
Los registros fósiles sugieren que las células procariotas aparecieron en la Tierra hace entre 3.500 y 3.800 millones de años. Todas las células están encerradas por una membrana celular, y el primer paso en la formación de las células sería la formación de esta membrana. Por lo tanto, vamos a investigar si una membrana celular podría formarse espontáneamente en condiciones prebióticas.
- Formación de la membrana celular
La membrana celular no es una estructura simple, sino compleja y dinámica, compuesta por lípidos (fosfolípidos, colesterol y glicolípidos), proteínas y carbohidratos. Los fosfolípidos forman la estructura bicapa fundamental, el colesterol modula la fluidez y los glicolípidos contribuyen al reconocimiento celular. Las proteínas, tanto integrales como periféricas, facilitan el transporte, la señalización y el soporte estructural, mientras que los carbohidratos desempeñan papeles cruciales en el reconocimiento y la comunicación celular. Esta composición permite a la membrana celular desempeñar sus funciones esenciales, manteniendo la homeostasis y facilitando las interacciones con el entorno.
La formación de una membrana celular por azar en condiciones prebióticas se enfrenta a varios problemas debido a la complejidad y especificidad que requieren las estructuras de membrana funcionales.
Las moléculas lipídicas anfifílicas específicas, como los fosfolípidos, requieren una combinación precisa de ácidos grasos, glicerol y grupos fosfato, que es poco probable que se formen y ensamblen espontáneamente en las proporciones correctas en condiciones prebióticas. La formación espontánea del grupo fosfato, como se ha demostrado en la sección anterior, es poco probable. Aunque las moléculas anfifílicas pueden formar bicapas espontáneamente, conseguir una bicapa estable y semipermeable capaz de encapsular y proteger un entorno celular requiere condiciones específicas. La aparición aleatoria de estas condiciones, incluida la concentración y los tipos de lípidos adecuados, es muy improbable.
El tamaño típico de una célula procariota, como una célula bacteriana, es de 1 micrómetro. Su superficie es de 3x10-12 m2 y el tamaño de una sola molécula de fosfolípido es de aproximadamente 5x10-19 m2. Por tanto, el número total de fosfolípidos en la bicapa es de 1,2x107. Para formar bicapas, aproximadamente diez millones de fosfolípidos deben alinearse uno al lado del otro y crear una cámara cerrada. Es muy poco probable que esto ocurra por azar, ya que las bicapas no se alinearían de forma natural y formarían una cámara cerrada sin algún tipo de guía o dirección.
Las condiciones de la Tierra primitiva eran duras y variables, con temperaturas extremas, niveles de pH y radiación. Mantener la integridad y estabilidad de una membrana primitiva en un entorno así habría sido todo un reto, ya que las membranas pueden verse fácilmente alteradas por estos factores. Una membrana funcional debe permitir de forma selectiva el paso de nutrientes y moléculas esenciales y mantener alejadas las sustancias nocivas. Esta permeabilidad selectiva requiere la presencia de proteínas y canales complejos, que difícilmente pueden formarse e integrarse en la membrana por procesos aleatorios.
Aunque se formaran membranas primitivas, la encapsulación aleatoria de las biomoléculas necesarias, como nucleótidos, aminoácidos y moléculas catalíticas, sería improbable. Es improbable que las concentraciones y combinaciones específicas necesarias para iniciar los procesos metabólicos primitivos se produzcan por azar.
La formación de una membrana funcional debe ir acompañada del desarrollo simultáneo de otra maquinaria celular, como proteínas de transporte y enzimas metabólicas, lo que complica aún más el escenario de formación de membranas a partir de procesos aleatorios. Así pues, la formación de células procariotas en la Tierra prebiótica no es factible.
vi. La formación de las células eucariotas
La teoría ampliamente aceptada sobre el origen de las células eucariotas es la teoría endosimbiótica. La teoría endosimbiótica sugiere que las células eucariotas se originaron a través de una relación simbiótica entre células procariotas primitivas. Este proceso consistió en el engullimiento de ciertas células procariotas (mitocondrias en el caso de las células animales y cloroplastos en el caso de las células vegetales) por una célula huésped ancestral, lo que dio lugar a una relación mutuamente beneficiosa y, finalmente, al desarrollo de células eucariotas complejas. Se afirma que la célula hospedadora ancestral era una arquea, pero los problemas de esta hipótesis son que la endocitosis, el proceso de engullir células procariotas, nunca se ha observado en las arqueas, y que la membrana celular de las arqueas está compuesta de enlaces éter, mientras que la membrana celular de las células eucariotas está compuesta de enlaces éster.
Esta teoría requiere células procariotas preexistentes y mitocondrias o cloroplastos. Sin embargo, el origen de las mitocondrias y los cloroplastos no está bien documentado. Las mitocondrias son orgánulos complejos con una estructura única que refleja su papel como centrales energéticas de la célula, generando ATP mediante la fosforilación oxidativa. Las mitocondrias están formadas por varios componentes distintos: la membrana externa, el espacio intermembranoso, la membrana interna y la matriz, que incluye enzimas, ADN, ribosomas y metabolitos. La membrana externa, al igual que la membrana celular, contiene una bicapa de fosfolípidos con una mezcla de fosfolípidos y proteínas. Es improbable que una estructura tan compleja pueda surgir espontáneamente a través de procesos aleatorios, ya que las membranas celulares, el ADN y las proteínas no pueden formarse espontáneamente. Las mitocondrias tienen su propio ADN, distinto del ADN nuclear, pero deben coordinarse con el genoma nuclear para funcionar correctamente. La integración del ADN mitocondrial en las redes reguladoras y metabólicas de una célula huésped plantea importantes retos.
El núcleo de las células eucariotas está compuesto por una membrana nuclear de doble capa, nucléolos y cromosomas, que contienen el material genético de la célula, incluidos el ADN, el ARN y las proteínas asociadas. El origen del núcleo en las células eucariotas es aún más difícil de explicar. Empecemos por el aspecto más sencillo: la membrana nuclear. El origen de la membrana nuclear en las células eucariotas es objeto de un importante debate científico. Se han propuesto varias hipótesis, entre ellas la de la invaginación de la membrana (plegamiento hacia dentro), la del origen vírico y la de la transferencia de genes, para explicar cómo pudo surgir esta compleja estructura.
La hipótesis de la invaginación de la membrana sugiere que la membrana nuclear se originó a partir de la invaginación de la membrana celular de una célula procariota ancestral. Sin embargo, esta hipótesis no explica la diferencia entre la membrana celular y la membrana nuclear. La membrana celular está compuesta por una única bicapa de fosfolípidos, mientras que la membrana nuclear consta de dos bicapas de fosfolípidos: una membrana interna y otra externa. Además, la membrana nuclear contiene complejos de poros nucleares que no se encuentran en la membrana celular. Además, la composición proteica de la membrana celular y de la membrana nuclear es diferente.
La hipótesis del origen vírico postula que los virus que infectaron las células primitivas podrían haber aportado material genético o componentes estructurales que acabaron dando lugar al desarrollo de una envoltura nuclear. La interacción entre las membranas virales y las de la célula huésped podría haber creado una estructura protectora alrededor del ADN. Aunque se sabe que los virus influyen en las estructuras de las células huésped, las pruebas concretas que vinculan a los virus con el origen de la membrana nuclear son limitadas.
La hipótesis de la transferencia de genes sugiere que la mezcla y transferencia de genes entre diferentes procariotas podría haber creado un genoma grande y complejo que requería un compartimento protector. La membrana nuclear habría evolucionado para proteger y regular este complejo material genético. Esta hipótesis se enfrenta a muchos problemas debido a la falta de pruebas directas, a su incapacidad para explicar cómo una estructura tan compleja y organizada de doble membrana y complejos de poros nucleares podría surgir únicamente de la transferencia e integración de genes, y a su incapacidad para proporcionar una vía clara de cómo los genes transferidos se integrarían y expresarían de forma que dieran lugar al desarrollo de la membrana nuclear.
La estructura de los nucleolos y los cromosomas es mucho más compleja que la de la membrana nuclear, por lo que resulta difícil imaginar que puedan originarse a partir de acontecimientos aleatorios. Además, es difícil comprender cómo estos componentes quedaron encerrados en la membrana. Los nucléolos y los cromosomas contienen la información genética de los organismos vivos, incluidos los planos para formar el ARN, las proteínas, el ADN, los orgánulos celulares y los tejidos y órganos de los seres vivos. El hecho de que estos planos para construir la vida estén previstos y ya presentes en el núcleo en la fase de célula eucariota, incluso antes de la formación de la vida, no puede ser explicado adecuadamente por la teoría evolucionista. Por el contrario, constituye una prueba evidente del diseño inteligente de la vida.
En resumen, el diseño inteligente puede explicar de forma natural el origen de las células eucariotas, mientras que la teoría de la evolución carece de una explicación clara de su origen.
vii. Localización de orgánulos
Las células están compuestas por varios orgánulos, como el núcleo, las mitocondrias, el retículo endoplásmico, el aparato de Golgi, los lisosomas y otros orgánulos, que trabajan conjuntamente para mantener la función y la homeostasis celulares. La localización de los orgánulos celulares es un proceso altamente regulado y dinámico que garantiza que los orgánulos se sitúen en una posición óptima dentro de la célula para mantener una función celular eficiente. Una localización adecuada es esencial para la salud celular y desempeña un papel fundamental en la adaptación a condiciones celulares y ambientales cambiantes. Cabe preguntarse cómo encuentran estos orgánulos su ubicación óptima, dado que no pueden pensar por sí mismos.

Fig. 3.4. Estructura de la célula animal y de la célula vegetal
Un examen detallado del proceso de localización de orgánulos revela un mecanismo muy preciso e intrincado que no puede atribuirse al azar. Este proceso implica una compleja interacción del citoesqueleto, las proteínas motoras, el tráfico de membranas, las proteínas de anclaje, los andamiajes, los ajustes dinámicos y la comunicación entre orgánulos.
El citoesqueleto desempeña un papel crucial en la localización de orgánulos. Proporciona soporte estructural, facilita el movimiento y garantiza la correcta posición de los orgánulos. El citoesqueleto se compone de tres tipos principales de filamentos: microtúbulos, filamentos de actina y filamentos intermedios, cada uno de los cuales contribuye de forma única a la localización de orgánulos.
Los microtúbulos son tubos largos y huecos formados por proteínas tubulinas. Forman una red que se extiende desde el centro organizador de microtúbulos (centrosoma) hasta la periferia celular. Los microtúbulos sirven de vías para las proteínas motoras, como la kinesina y la dineína. La kinesina desplaza los orgánulos hacia el extremo positivo de los microtúbulos, normalmente hacia la periferia celular, mientras que la dineína los desplaza hacia el extremo negativo, normalmente hacia el centro celular. Los microtúbulos ayudan a posicionar orgánulos como el aparato de Golgi, que suele estar situado cerca del centrosoma, y las mitocondrias, que están distribuidas por toda la célula pero pueden ser transportadas a lo largo de los microtúbulos de a zonas con gran demanda de energía.

Fig. 3.5. los microtúbulos y las proteínas motoras
Los filamentos de actina, también conocidos como microfilamentos, son fibras finas y flexibles formadas por la proteína actina. Se concentran justo debajo de la membrana plasmática y forman una densa red por todo el citoplasma. Los filamentos de actina facilitan el flujo citoplasmático, un proceso que ayuda a distribuir orgánulos y nutrientes por toda la célula. Las proteínas motoras de la miosina interactúan con los filamentos de actina para transportar vesículas, endosomas y otros pequeños orgánulos a lo largo de la red de actina. Los filamentos de actina ayudan a mantener la forma celular y participan en el movimiento celular, que afecta indirectamente a la posición de los orgánulos.
Los filamentos intermedios son fibras en forma de cuerda formadas por diversas proteínas (como queratinas, vimentina y láminas) en función del tipo de célula. Proporcionan resistencia mecánica y soporte estructural. Los filamentos intermedios ayudan a estabilizar la posición de orgánulos como el núcleo, anclándolos en su lugar dentro del citoplasma. Mantienen la integridad general del citoesqueleto, garantizando que otros componentes como los microtúbulos y los filamentos de actina puedan funcionar eficazmente en la localización de orgánulos.
Los diferentes tipos de filamentos del citoesqueleto a menudo trabajan juntos para posicionar los orgánulos con precisión. Por ejemplo, los microtúbulos y los filamentos de actina se coordinan para garantizar la distribución y el movimiento adecuados de vesículas y orgánulos. El citoesqueleto es muy dinámico y se remodela continuamente para adaptarse a las necesidades de la célula. Esta flexibilidad permite el rápido reposicionamiento de orgánulos en respuesta a señales celulares o cambios en el entorno.
El tráfico de membranas es el proceso por el que proteínas, lípidos y otras moléculas se transportan dentro de las células, garantizando que los componentes celulares lleguen a sus destinos correctos. Para ello, las vesículas brotan de las membranas donantes, se transportan por el citoplasma y se fusionan con las membranas de destino. Los principales orgánulos implicados en el tráfico de membranas son el retículo endoplásmico, el aparato de Golgi y varios tipos de vesículas como endosomas y lisosomas. Este proceso es esencial para mantener la organización celular, facilitar la comunicación entre orgánulos y permitir que la célula responda eficazmente a señales internas y externas.
Las vías de señalización guían el movimiento y la posición de los orgánulos dentro de la célula. Estas vías implican la transmisión de señales químicas que proporcionan indicaciones espaciales, garantizando que los orgánulos se dirijan a sus ubicaciones apropiadas. Los receptores de las superficies de los orgánulos y del citoplasma interactúan con las moléculas señalizadoras para facilitar este proceso. Por ejemplo, las pequeñas GTPasas como las proteínas Rab son reguladores clave que controlan el tráfico de vesículas y el posicionamiento de orgánulos mediante la interacción con proteínas efectoras específicas. Estas vías de señalización garantizan la coordinación de los procesos celulares y el posicionamiento dinámico de los orgánulos en respuesta a los cambios de las necesidades celulares y las condiciones ambientales.
Las proteínas de anclaje y los andamiajes desempeñan un papel vital en la localización celular, ya que garantizan que los orgánulos se sitúen con precisión dentro de la célula. Las proteínas de anclaje conectan orgánulos a lugares específicos del citoplasma, estabilizándolos e impidiendo su desplazamiento. Por ejemplo, las mitocondrias pueden fijarse al retículo endoplásmico mediante mecanismos de anclaje específicos, lo que facilita la transferencia eficiente de energía y la coordinación metabólica. Las proteínas de andamiaje proporcionan soporte estructural formando complejos que sujetan los orgánulos en su sitio, manteniendo la organización general de la célula. Estas proteínas crean un marco dinámico que permite la correcta disposición de los orgánulos, garantizando que las funciones celulares se lleven a cabo con eficacia y eficiencia.
Los ajustes dinámicos en la localización celular se refieren a los cambios continuos y sensibles en el posicionamiento de los orgánulos dentro de una célula. Estos ajustes son cruciales para mantener la función y adaptabilidad celulares. Durante las distintas fases del ciclo celular, como la mitosis, orgánulos como el núcleo y las mitocondrias se reposicionan para garantizar la correcta división celular. Además, en respuesta a estímulos ambientales, como la disponibilidad de nutrientes o las condiciones de estrés, los orgánulos pueden reubicarse en zonas donde sus funciones sean más necesarias. Esta reubicación dinámica se ve facilitada por el citoesqueleto y las proteínas motoras, lo que permite a la célula mantener la homeostasis y responder eficazmente a las cambiantes condiciones internas y externas.
La comunicación entre orgánulos garantiza la coordinación y la eficacia de las funciones celulares. Esta comunicación se produce a través de sitios de contacto directo y transporte vesicular. Los puntos de contacto, como las membranas asociadas a las mitocondrias (MAM) entre las mitocondrias y el retículo endoplásmico, facilitan la transferencia de lípidos, calcio y otras moléculas, garantizando la sincronización de las actividades entre orgánulos. El transporte vesicular implica el desprendimiento y la fusión de vesículas, que transportan proteínas y lípidos entre orgánulos, manteniendo su integración funcional. La comunicación eficaz entre orgánulos es esencial para procesos como el metabolismo, la señalización y las respuestas al estrés, contribuyendo a la homeostasis general de la célula.
Como se ha descrito anteriormente, los mecanismos implicados en la localización de orgánulos son muy organizados y complejos. La evolución paso a paso de sistemas tan intrincadamente coordinados mediante mutaciones aleatorias y selección natural es extremadamente improbable por las siguientes razones.
No existen pruebas directas de etapas intermedias en la evolución de los mecanismos de localización de orgánulos. Los registros fósiles y los estudios moleculares no captan las formas de transición que ilustrarían la evolución gradual de estos sofisticados sistemas. La complejidad de la localización de orgánulos y su coordinación dentro de las células plantea un reto a las explicaciones evolutivas, ya que la organización celular presenta una ‘complejidad irreducible’, en la que la eliminación de cualquier parte haría que el sistema dejara de ser funcional. La teoría evolutiva explica la complejidad mediante modificaciones graduales, pero las estructuras celulares y su localización precisa no tienen etapas intermedias viables.
La localización de orgánulos depende de intrincadas interacciones con el citoesqueleto, las proteínas motoras, las vías de señalización y otros componentes celulares. Esta interdependencia plantea interrogantes sobre cómo han podido coevolucionar estos sistemas de forma escalonada. Resulta difícil explicar cómo tanto los orgánulos como los sistemas responsables de su localización pudieron evolucionar simultáneamente sin que uno de ellos fuera plenamente funcional en primer lugar.
El origen y la evolución de proteínas motoras como la kinesina, la dineina y la miosina, así como de elementos citoesqueléticos como los microtúbulos y los filamentos de actina, no se conocen del todo. Estas proteínas y estructuras deben haber desarrollado funciones e interacciones muy específicas, difíciles de explicar únicamente mediante cambios incrementales. La evolución de las complejas redes reguladoras que controlan la localización de orgánulos plantea importantes retos. Estas redes deben coordinar con precisión la expresión y actividad de numerosos genes, y su evolución incremental a través de mutaciones aleatorias es difícil de explicar.
Muchos componentes implicados en la localización de orgánulos son interdependientes, lo que significa que deben funcionar juntos de forma eficaz para proporcionar alguna ventaja selectiva. La evolución simultánea de múltiples partes que interactúan es problemática porque los sistemas parciales no conferirán un beneficio suficiente para ser favorecidos por la selección natural.
Los procesos de localización y mantenimiento de orgánulos consumen mucha energía. No está claro cómo las células primitivas podían permitirse los costes metabólicos asociados a estos complejos sistemas sin disponer ya de mecanismos eficientes de producción de energía y gestión de recursos.
viii. Diferenciación celular
La diferenciación celular es el proceso por el cual células no especializadas se convierten en células especializadas con estructuras y funciones distintas.Este proceso es crucial para el desarrollo, crecimiento y funcionamiento de tejidos, órganos y, en última instancia, organismos pluricelulares. La diferenciación suele comenzar con las células madre, que son células indiferenciadas capaces de dar lugar a diversos tipos celulares. Las células madre pueden ser pluripotentes, capaces de diferenciarse en casi cualquier tipo celular. Durante el desarrollo, estas células reciben señales que las guían para convertirse en tipos celulares específicos. A medida que las células madre se diferencian, se convierten en células progenitoras multipotentes, que se comprometen a dar lugar a una gama limitada de tipos celulares. Las células progenitoras se diferencian a su vez en células totalmente especializadas. La diferenciación celular es un proceso altamente regulado y dinámico que depende de la regulación de la expresión génica, las vías de transducción de señales, las modificaciones epigenéticas, los gradientes morfogénicos y las interacciones con otras células y la matriz extracelular.
Todas las células de un organismo contienen el mismo ADN, pero los distintos tipos celulares expresan diferentes subconjuntos de genes. Esta expresión génica selectiva impulsa la diferenciación. Las proteínas conocidas como factores de transcripción se unen a secuencias específicas de ADN para regular la transcripción de genes diana. Estos factores pueden activar o reprimir la expresión génica, dando lugar a la producción de proteínas necesarias para un tipo celular específico.
Las células reciben señales de su entorno, como factores de crecimiento, hormonas y citoquinas. Estas señales se unen a receptores de la superficie celular, iniciando vías de transducción de señales. Las vías de transducción de señales implican una cascada de acontecimientos intracelulares, que a menudo incluyen la fosforilación de proteínas, que en última instancia dan lugar a cambios en la expresión génica.
Las modificaciones epigenéticas implican la metilación del ADN y la modificación de las histonas. La metilación del ADN silencia la expresión génica añadiendo grupos metilo al ADN, normalmente en islas CpG. Los patrones de metilación son hereditarios y pueden fijar la identidad de una célula reprimiendo genes que no son necesarios para un tipo celular concreto. Las histonas, las proteínas alrededor de las cuales se enrolla el ADN, pueden modificarse químicamente (por ejemplo, acetilación, metilación). Estas modificaciones alteran la estructura de la cromatina, haciendo que el ADN sea accesible para la transcripción.
Los morfógenos son moléculas señalizadoras que se difunden por los tejidos y forman gradientes de concentración. Las células responden a diferentes concentraciones de morfógenos activando distintas vías de desarrollo, lo que conduce a diversos destinos celulares. Los gradientes de morfógenos son cruciales en el desarrollo embrionario para la formación de patrones, determinando la disposición espacial de las células diferenciadas.
El contacto directo entre células puede inducir la diferenciación. Las proteínas de membrana de una célula interactúan con las proteínas receptoras de una célula adyacente para transmitir señales. Las células segregan moléculas de señalización que afectan a las células cercanas, influyendo en su diferenciación.
La matriz extracelular (MEC), compuesta por proteínas y polisacáridos, proporciona soporte estructural y señales bioquímicas a las células. Las integrinas y otras moléculas de adhesión median en la unión de las células a la MEC, influyendo en la forma, migración y diferenciación celular.
Los mecanismos de retroalimentación positiva y negativa controlan el progreso de la diferenciación. La retroalimentación positiva indica que las células diferenciadas pueden producir señales que refuerzan su identidad, asegurando tipos celulares estables. Los mecanismos de retroalimentación negativa limitan las señales de diferenciación, impidiendo una diferenciación excesiva y manteniendo una reserva de células indiferenciadas.
Como se ha descrito, la diferenciación celular implica una serie de acontecimientos muy complejos y coordinados, que incluyen una regulación génica precisa, la transducción de señales y modificaciones epigenéticas. Esta complejidad es difícil de explicar únicamente a través de mutaciones graduales y aleatorias y de la selección natural. El proceso requiere la integración de numerosos sistemas celulares, como los factores de transcripción, las vías de señalización y el citoesqueleto. La evolución simultánea de estos sistemas interdependientes plantea un reto importante a la teoría evolutiva. Además, el origen de las células madre pluripotentes no puede explicarse mediante mecanismos evolutivos.
El papel de las modificaciones epigenéticas, como la metilación del ADN y la modificación de las histonas, es crucial en la diferenciación. El origen de estos sofisticados mecanismos no está bien explicado por la teoría evolutiva, ya que requieren un alto nivel de precisión y coordinación. La heredabilidad de las marcas epigenéticas añade otra capa de complejidad. Los mecanismos por los que estas marcas se establecen, mantienen y heredan son intrincados y requieren una explicación detallada.
El establecimiento y la interpretación de gradientes de morfógenos son fundamentales para la formación de patrones durante el desarrollo. Los gradientes de concentración precisos y la capacidad de la célula para interpretar con exactitud estas señales sugieren un diseño inteligente y no mutaciones aleatorias. El concepto de información posicional, por el que las células determinan su ubicación y se diferencian en consecuencia, requiere un sofisticado sistema de comunicación. El origen evolutivo de dicho sistema no se conoce con claridad.
Las redes reguladoras de los factores de transcripción que controlan la expresión génica durante la diferenciación son muy complejas. La evolución incremental de estas redes carece de apoyo empírico, dada la necesidad de cambios coordinados en múltiples genes. Las mutaciones en factores de transcripción clave pueden tener efectos generalizados y deletéreos, por lo que resulta difícil imaginar cómo podrían acumularse gradualmente mutaciones beneficiosas para formar redes reguladoras funcionales.
ix. La formación de tejidos y órganos
La formación de tejidos (histogénesis) es el proceso por el cual las células diferenciadas se organizan en tejidos específicos durante el desarrollo embrionario.
Este proceso implica la especialización de las células madre en varios tipos celulares, como células musculares, nerviosas y epiteliales, cada una con funciones distintas. Una vez que las células se diferencian, empiezan a organizarse en estructuras complejas que forman los tejidos básicos del cuerpo. Estos tejidos incluyen el epitelial, el conjuntivo, el muscular y el nervioso, cada uno de los cuales contribuye a la estructura y función general de los órganos.
La comunicación celular y las vías de señalización desempeñan un papel crucial a la hora de guiar a las células a sus ubicaciones correctas y garantizar que interactúen adecuadamente. La histogénesis está estrechamente regulada, ya que los errores en la organización celular pueden provocar anomalías en el desarrollo o enfermedades. A lo largo de este proceso, las células se adhieren unas a otras, migran a regiones específicas y sufren cambios morfológicos para formar estructuras tisulares funcionales. La finalización de la histogénesis da lugar a la formación de tejidos plenamente desarrollados capaces de desempeñar funciones especializadas. Este proceso es fundamental para el correcto desarrollo de los órganos y la organización general del cuerpo.
La formación de órganos (organogénesis) sigue a la histogénesis, en la que los tejidos se organizan en unidades funcionales. Durante la organogénesis, las tres capas germinales (ectodermo, mesodermo y endodermo) interactúan y se diferencian para formar órganos específicos. El ectodermo forma principalmente órganos como el cerebro y la médula espinal, mientras que el mesodermo da lugar al corazón, los riñones y los músculos esqueléticos. El endodermo forma estructuras internas como los pulmones y el hígado.
La organogénesis implica complejas vías de señalización y regulación genética para garantizar que los órganos se desarrollen en la ubicación correcta y con la función adecuada. Durante la organogénesis, las células migran, proliferan y se someten a apoptosis según sea necesario para dar forma a los órganos en desarrollo. La vía de señalización Notch es especialmente importante para determinar el destino celular y mantener el equilibrio entre proliferación y diferenciación celular. La señalización Wnt contribuye al patrón y la morfogénesis de los órganos, garantizando que los tejidos se desarrollen en las ubicaciones y proporciones correctas. Las alteraciones de esta señalización pueden provocar defectos congénitos o un desarrollo anormal de los órganos. Este proceso es crucial para establecer la anatomía y fisiología general del cuerpo.
A medida que los órganos se desarrollan, múltiples tipos de tejido se integran y funcionan juntos. Por ejemplo, un órgano como el corazón se compone de tejido muscular, tejido conjuntivo y tejido nervioso, todos ellos esenciales para su funcionamiento. El desarrollo de estos órganos está guiado por complejas vías de señalización que garantizan que las células migren a los lugares correctos, se diferencien adecuadamente y formen las estructuras correctas.
Las teorías evolutivas que explican la formación de tejidos y órganos se enfrentan a importantes desafíos. La complejidad de los tejidos y órganos es demasiado grande para explicarla mediante procesos evolutivos graduales, paso a paso. Muchos tejidos y órganos presentan una ‘complejidad irreducible’, es decir, constan de múltiples partes interdependientes que no podrían funcionar si faltara alguna de ellas. Estas estructuras complejas no podrían haber evolucionado de forma gradual, ya que no serían funcionales en etapas intermedias.
La teoría evolutiva postula que las nuevas estructuras, como los tejidos y los órganos, surgen mediante la modificación gradual de las estructuras existentes. Sin embargo, esto no explica adecuadamente el origen de estructuras totalmente nuevas que no tienen precursores aparentes. Por ejemplo, el desarrollo de órganos complejos como el cerebro o el sistema inmunitario se considera difícil de explicar mediante pequeños cambios graduales.
La información genética necesaria para construir y organizar tejidos y órganos es vasta y muy específica, y es poco probable que una información tan detallada surja a través de mutaciones aleatorias.
Los factores epigenéticos, que influyen en la expresión de los genes sin modificar la secuencia del ADN, desempeñan un papel importante en el desarrollo de tejidos y órganos. La teoría de la evolución, que hace hincapié en las mutaciones genéticas, no tiene plenamente en cuenta la complejidad añadida que introduce la regulación epigenética. También se queda corta a la hora de explicar cómo sistemas biológicos complejos (compuestos por múltiples tejidos y órganos que interactúan entre sí) pueden evolucionar de forma independiente e integrarse posteriormente en para funcionar de forma cohesionada como un organismo unificado.
x. La formación de los organismos multicelulares
Una vez formados los órganos individuales de, deben integrarse en un organismo cohesionado y funcional. Esta integración se consigue mediante la organización espacial de los órganos dentro del cuerpo, donde cada órgano ocupa una ubicación específica que le permite interactuar con otros órganos y sistemas. Por ejemplo, el sistema circulatorio, que incluye el corazón y los vasos sanguíneos, debe estar correctamente conectado con otros sistemas, como el respiratorio y el digestivo, para mantener la vida.
A lo largo de este proceso, las células de los tejidos y órganos siguen especializándose y adaptándose a sus funciones, un proceso conocido como diferenciación funcional. Esto garantiza que cada parte del organismo desempeñe eficazmente las funciones que le han sido asignadas. La coordinación y la interacción entre los distintos órganos y sistemas son esenciales para mantener la salud y el funcionamiento general del organismo pluricelular, permitiéndole sobrevivir, crecer y reproducirse. La explicación evolutiva de la formación de organismos pluricelulares a partir de órganos implica abordar varios retos y complejidades clave:
La formación de organismos pluricelulares a partir de órganos requiere un nivel increíblemente alto de integración y coordinación entre varios sistemas. Los procesos evolutivos que podrían conducir al desarrollo simultáneo y al funcionamiento sin fisuras de múltiples sistemas de órganos son difíciles de explicar.
Los órganos y sistemas de los organismos pluricelulares son muy interdependientes, lo que significa que la funcionalidad de un sistema depende a menudo del buen funcionamiento de otros. Las explicaciones evolutivas deben dar cuenta del desarrollo simultáneo de distintos órganos y sistemas, cada uno con funciones e interdependencias específicas, y explicar cómo evolucionaron estos sistemas complejos de forma coordinada y paso a paso. Las formas intermedias con sistemas parcialmente desarrollados no aportarían ventajas suficientes para ser favorecidas por la selección natural.
En el registro fósil escasean las formas de transición claras que ilustren la evolución gradual de organismos pluricelulares simples a organismos complejos con órganos completamente formados. Esta laguna dificulta el rastreo de las vías evolutivas que condujeron al desarrollo de estructuras tan complejas.
La coordinación precisa de la expresión génica y las vías de desarrollo necesarias para la formación e integración de órganos plantea importantes retos. Pequeños errores en estos procesos pueden dar lugar a trastornos del desarrollo, lo que plantea interrogantes sobre cómo podrían evolucionar de forma incremental sistemas tan delicados.
El desarrollo de organismos multicelulares complejos requiere mecanismos robustos para gestionar errores y variaciones. La explicación evolutiva debe explicar cómo evolucionaron estos sistemas de gestión de errores y cómo garantizan la estabilidad y fidelidad de la formación y función de los órganos.
b. ¿Puede la evolución explicar el origen de la vida?
En la sección anterior hemos tratado el origen de la vida, trazando su progresión desde la formación de aminoácidos, ARN, proteínas, ADN, células procariotas, células eucariotas, tejidos y órganos, hasta llegar a los organismos pluricelulares. Es innegable que estos procesos han progresado en de una manera dirigida y guiada hacia un propósito singular: la formación de organismos vivos.
Esto plantea una cuestión importante: ¿Puede la evolución, que funciona mediante procesos no dirigidos y aleatorios, explicar adecuadamente estos complejos desarrollos y el origen de la vida? Los científicos evolucionistas han propuesto diversas teorías para responder a esta pregunta. Las principales teorías de la evolución son la selección natural, la mutación, la deriva genética y la transferencia horizontal de genes. Veamos brevemente cada una de estas teorías.
La selección natural es el proceso por el cual los individuos con rasgos ventajosos sobreviven y se reproducen con más éxito, lo que hace que esos rasgos se vuelvan más comunes en una población a lo largo de generaciones. La selección natural actúa sobre las variaciones existentes en los organismos vivos. Así pues, el origen de la vida y la formación de sus componentes fundamentales (aminoácidos, ARN, proteínas, ADN) y estructuras (células, tejidos, órganos y organismos pluricelulares) requieren explicaciones que van más allá de la selección natural, ya que estos procesos carecen de las condiciones previas necesarias (replicación y funcionalidad) para que actúe la selección.
Las mutaciones son cambios aleatorios en el ADN de un organismo que pueden introducir variaciones genéticas, a veces dando lugar a nuevos rasgos o adaptaciones. La mutación plantea problemas porque la mayoría de las mutaciones son perjudiciales o neutras, más que beneficiosas, lo que hace improbable que las mutaciones ventajosas se produzcan con la frecuencia suficiente para impulsar un cambio evolutivo significativo. Por ejemplo, un estudio sobre la distribución de los efectos sobre la aptitud (DFE) de las mutaciones aleatorias en el virus de la estomatitis vesicular ilustra este problema. De todas las mutaciones, el 39,6% eran letales, el 31,2% deletéreas no letales y el 27,1% neutras.

Fig. 3.6. Distribución del efecto de aptitud
Si se insertan o eliminan nucleótidos (causando mutaciones de desplazamiento de marco), o si se crean o eliminan codones de parada mediante mutaciones, se producen proteínas no funcionales. Ésta es una de las principales razones por las que, teniendo en cuenta el gran número de aminoácidos que contienen las proteínas de los organismos vivos (por ejemplo, de 20 a 33.000 en las proteínas humanas), la probabilidad de que se produzca una macroevolución a través de este tipo de mutaciones aleatorias es imposible (véase el apartado ‘d’ de este capítulo para más detalles). Además, las mutaciones aleatorias no pueden explicar la aparición inicial de la vida a partir de materia inerte.
La deriva genética se basa en cambios aleatorios en las frecuencias alélicas, que pueden no explicar suficientemente la complejidad adaptativa observada en los organismos. La deriva genética es más pronunciada en poblaciones pequeñas, por lo que su impacto es menos relevante en poblaciones más grandes, donde se produce la mayor parte de la evolución. Además, carece de la fuerza direccional necesaria para explicar el desarrollo de estructuras y sistemas altamente organizados. Además, la deriva genética no puede producir información o funciones nuevas, por lo que no explica la aparición de nuevos rasgos ni el origen de características biológicas complejas en.
La transferencia horizontal de genes (HGT) es la transferencia de material genético entre organismos no relacionados, no a través de la herencia, lo que contribuye a la variación genética. La transferencia horizontal de genes plantea problemas a la hora de explicar rasgos complejos en organismos pluricelulares, ya que su papel se limita principalmente a los procariotas y tiene menos impacto en los organismos superiores. La integración de genes extraños en el genoma de un huésped requiere a menudo mecanismos reguladores precisos, que es poco probable que evolucionen simultáneamente. Además, el HGT puede introducir inestabilidad genética, lo que puede dar lugar a mutaciones perjudiciales. La naturaleza aleatoria de la adquisición de genes mediante HGT también plantea dudas sobre su capacidad para producir adaptaciones coordinadas y funcionales. El HGT no explica el origen de nuevos genes, sino más bien la transferencia de los ya existentes, por lo que no aborda la aparición de nuevos rasgos.
La siguiente tabla resume la aplicabilidad de las teorías evolutivas a la biogénesis y los procesos genéticos.
Teorías de la evolución | ¿Puede explicar la biogénesis? | ¿Puede explicar la formación del ARN, las proteínas y el ADN? | ¿Adaptación genética, no evolución?* |
Selección natural | No | No | Sí |
Mutación | No | No | Sí |
Deriva genética | No | No | Sí |
HGT | No | No | N/A |
Tabla 3.2. Teorías de la evolución: aplicabilidad a la biogénesis y la genética (*: véase la sección siguiente para la adaptación genética)
Como se muestra en la tabla, las principales teorías evolutivas no logran explicar el origen de la vida en la Tierra ni los mecanismos que subyacen a la formación de componentes biológicos fundamentales como el ARN, las proteínas y el ADN. Esto sugiere que los modelos evolutivos aplicados a células, tejidos, órganos y formas de vida existentes no constituyen verdaderas explicaciones del origen o la evolución de la vida en sí. En lugar de abordar el surgimiento de la vida a partir de materia no viva, estas teorías se limitan a describir cómo se desarrolla la vida una vez que los componentes esenciales -ARN, proteínas y ADN- ya están en su lugar, de forma parecida a detallar el proceso de ensamblaje de un coche o la construcción de un edificio sin explicar cómo llegaron a existir las materias primas y las piezas.
Las teorías evolutivas aplicadas a los organismos vivos describen principalmente los procesos genéticos y bioquímicos que les permiten adaptarse a entornos cambiantes. Sin embargo, estas adaptaciones y comportamientos no son creados por la evolución, sino que ya están codificados en su información genética. Dada esta limitación, las teorías evolutivas se denominarían más bien ‘Teoría de la adaptación genética’ (véase la siguiente sección), ya que abordan principalmente las formas en que los organismos se ajustan a las presiones ambientales a través de mecanismos genéticos preexistentes.
A pesar de estas limitaciones críticas, la teoría de la evolución se ha promovido excesivamente, creando conceptos erróneos generalizados. Mucha gente cree ahora erróneamente que puede explicar la transición de la materia no viva a los organismos vivos y el desarrollo de formas de vida complejas.
Para construir un edificio, necesitamos planos, materiales de construcción y unos cimientos sólidos. Las teorías evolucionistas son como intentar construir un edificio sin planos (direccionalidad), materiales de construcción (ARN, proteínas, ADN) y cimientos (el origen inicial de la vida). Sin ellos, no se pueden construir edificios.
Al igual que reconocemos que los planos de un edificio fueron diseñados por un arquitecto, también deberíamos reconocer que todos los organismos vivos fueron diseñados y creados por Dios, el Creador divino.
c. Teoría de Darwin: ¿Teoría de la Evolución o Teoría de la Adaptación Genética?
A grandes rasgos, la evolución se divide en dos tipos: microevolución y macroevolución. La microevolución se refiere a los cambios a pequeña escala que se producen en una especie a lo largo del tiempo. Estos cambios son observables a corto plazo y a menudo implican una adaptación al entorno. La macroevolución, en cambio, implica cambios a gran escala que se producen a lo largo de períodos geológicos prolongados y que dan lugar a la formación de nuevas especies y grupos taxonómicos más amplios.
Los biólogos evolutivos proponen que el principal mecanismo de la macroevolución es la acumulación de numerosos cambios microevolutivos a lo largo del tiempo. La gente está de acuerdo en que hay pruebas de microevolución, pero no pruebas convincentes de macroevolución. Para que el darwinismo pueda considerarse la teoría de la evolución, debe mostrar las pruebas de la macroevolución. La prueba más convincente de la macroevolución es la existencia de especies de transición. En el capítulo 6 (Dificultades para la teoría) del libro de Darwin ‘Sobre el origen de las especies’, está escrito: ‘¿por qué, si las especies han descendido de otras especies por graduaciones insensiblemente finas, no vemos por todas partes innumerables formas transicionales?’. Esta falta de pruebas de las especies transicionales se conoce a menudo como ‘el dilema de Darwin’.
Los fósiles a menudo etiquetados como ‘transicionales’ podrían ser simplemente variaciones dentro de una especie o formas no relacionadas entre sí. Esta ambigüedad dificulta la identificación concluyente de las verdaderas formas transicionales. Por ejemplo, Tiktaalik se considera un fósil transicional y uno de los descubrimientos más importantes en el estudio de la evolución de los vertebrados. Sin embargo, un artículo publicado en Nature por Niedzwiedzki et al. revela huellas de tetrápodos bien conservadas que son anteriores a Tiktaalik en unos 18 millones de años. Las huellas descubiertas sugieren que los tetrápodos plenamente desarrollados ya caminaban sobre tierra firme mucho antes de lo que se creía. Dado que Tiktaalik data de hace unos 375 millones de años, la presencia de huellas de tetrápodos más antiguas cuestiona su papel como forma de transición directa entre los peces y los tetrápodos.
Si no hay pruebas convincentes de la existencia de especies de transición, la teoría de Darwin se denominó erróneamente y debería llamarse ‘teoría de la adaptación genética’ en lugar de ‘teoría de la evolución’. La razón está relacionada con los ciclos de Milankovitch, que influyen en los patrones climáticos y han desempeñado un papel en la configuración de las adaptaciones genéticas a lo largo del tiempo.
- Ciclos Milankovitch
La excentricidad de la Tierra fluctúa de casi circular a más elíptica a lo largo de un ciclo de 100.000 años. El cambio de excentricidad influye en los patrones climáticos, contribuyendo al calendario de los periodos glaciares e interglaciares.
La inclinación axial de la Tierra (oblicuidad) varía entre 22,1 grados y 24,5 grados a lo largo de un ciclo de 41.000 años. Esta inclinación afecta a la distribución de la radiación solar entre el ecuador y los polos, influye en la intensidad de las estaciones y desempeña un papel crucial en los patrones climáticos a largo plazo y en la dinámica de la era glacial .
La precesión del eje de rotación de la Tierra implica el cambio gradual de la orientación del eje a lo largo de un ciclo de 26.000 años. Este bamboleo hace que el calendario de las estaciones cambie en relación con la posición de la Tierra en su órbita. Este mecanismo altera la intensidad y el calendario de las estaciones, lo que repercute en el sistema climático global de la Tierra.
Los efectos combinados de los cambios en la excentricidad, la inclinación axial y la precesión del eje de rotación se conocen colectivamente como ciclos de Milankovitch. Estos ciclos provocan cambios climáticos globales a largo plazo. El desierto del Sahara es un buen ejemplo de cambio climático. Durante los periodos de mayor radiación solar, el Sáhara experimenta más precipitaciones, lo que lo transforma en un paisaje verde y exuberante con lagos y ríos. Por el contrario, la disminución de la radiación solar provoca condiciones de aridez, convirtiendo la región en el vasto desierto que se ve hoy en día.

Fig. 3.7. Componentes de los ciclos de Milankovitch
Cuando se producen estos cambios, todos los organismos vivos de la Tierra ajustan sus cuerpos a los entornos cambiantes mediante la adaptación genética. Este extraordinario mecanismo, codificado en el ADN, permite a los organismos sobrevivir durante largos periodos de tiempo sin extinguirse. Aunque los evolucionistas han calificado tradicionalmente esta adaptabilidad como ‘evolución’, tal clasificación es engañosa; sería más preciso y científico describirla como ‘adaptación genética’. Permítanme ilustrar algunos ejemplos que podrían respaldar el concepto de la ‘teoría de la adaptación genética’.
- Adaptación genética a la radiación UV
Si la piel humana se expone a una fuerte radiación UV debido al cambio climático, un complejo mecanismo en el que intervienen varias proteínas y hormonas desencadena un aumento de la producción de melanina mediante la activación de genes específicos.

Fig. 3.8. Mecanismo de producción de melanina
La radiación UV provoca daños en el ADN de las células de la piel. Este daño activa la proteína p53, que es un regulador crucial de la respuesta de la célula al estrés y al daño. La proteína p53 activada actúa como factor de transcripción, promoviendo la expresión de varios genes implicados en la respuesta protectora al daño UV. P53 estimula la expresión del gen de la pro-opiomelanocortina (POMC). La POMC es un polipéptido precursor que puede escindirse en varios péptidos más pequeños con diferentes funciones. La POMC se transforma en múltiples péptidos, como la hormona adrenocorticotrópica (ACTH) y la hormona estimulante de melanocitos (MSH).
La MSH se une al receptor de melanocortina 1 (MC1R) en la superficie de los melanocitos, las células responsables de la producción de melanina. La unión de la MSH al MC1R activa el receptor, lo que desencadena una cascada de señales en el interior de los melanocitos. La activación del MC1R conduce a la regulación al alza de los genes implicados en la síntesis de melanina. Los melanocitos aumentan la producción de melanina, un pigmento que absorbe y disipa la radiación UV, protegiendo así el ADN de las células de la piel de nuevos daños inducidos por los rayos UV.
La melanina se empaqueta en melanosomas, que luego se transportan a los queratinocitos, el tipo de célula predominante en la capa externa de la piel. La melanina forma una capa protectora sobre los núcleos de los queratinocitos, protegiendo eficazmente el ADN de la radiación UV.
Este es uno de los ejemplos de adaptación genética en respuesta a un entorno cambiante en un periodo de tiempo relativamente corto.
- Adaptación genética al entorno ártico
Los inuit han desarrollado adaptaciones genéticas que les permiten prosperar en el duro entorno ártico. Las principales adaptaciones de incluyen variantes en el grupo de genes de la desaturasa de ácidos grasos (FADS), que mejoran su capacidad para metabolizar los ácidos grasos omega-3 y omega-6 de su dieta tradicional rica en grasas de mamíferos marinos. Además, los cambios genéticos en el gen de la carnitina palmitoiltransferasa 1A (CPT1A) mejoran la producción de energía a partir de las grasas, crucial para mantener el calor corporal. Estas adaptaciones reducen el riesgo de enfermedades cardiovasculares a pesar de una dieta rica en grasas. Además, la adaptación en los genes que regulan la actividad de la grasa parda mejora la termogénesis, ayudando a los inuit a generar calor y mantener la temperatura corporal en condiciones de frío extremo. En conjunto, estas adaptaciones genéticas favorecen su supervivencia en condiciones climáticas frías. Estos cambios parecen datar de hace al menos 20.000 años, cuando los antepasados inuit vivían alrededor del estrecho de Bering, entre Rusia y Alaska. Se trata de otro ejemplo de adaptación genética a un entorno cambiante.

Fig. 3.9. Inuit cuyos genes se adaptaron al ambiente frío
- De oso pardo a oso polar por adaptación genética
La transición del oso pardo al oso polar es un buen ejemplo de adaptación genética impulsada por presiones ambientales.Hace aproximadamente 400.000 años, una población de osos pardos quedó aislada en el Ártico, donde se enfrentó a diferentes retos de supervivencia. Con el tiempo, se seleccionaron de forma natural los cambios genéticos que les conferían ventajas en el duro y gélido entorno.

Fig. 3.10. Oso pardo y oso polar
Las adaptaciones clave incluyen cambios en genes relacionados con el metabolismo de las grasas, como el gen de la apolipoproteína B (APOB), que mejoró la capacidad de procesar una dieta rica en grasas de las focas, su principal fuente de alimento. Las adaptaciones en genes como el receptor de endotelina tipo B (EDNRB) y el ausente en el melanoma 1 (AIM1) también propiciaron el desarrollo del pelaje blanco, que les proporciona camuflaje contra la nieve y el hielo. Además, los cambios genéticos que afectan a la estructura esquelética y la morfología de las extremidades del oso mejoraron su capacidad natatoria, crucial para cazar en aguas árticas.
Estas adaptaciones genéticas permitieron a los osos polares explotar eficazmente los recursos del Ártico, sobrevivir en el frío extremo y diferenciarse de sus antepasados los osos pardos. Es importante señalar que, a pesar de 400.000 años de cambios genéticos, siguen siendo osos y no se han transformado en una especie diferente.
- Cambio de pico en los pinzones por adaptación genética
El cambio de tamaño y forma del pico en los pinzones de Darwin es un ejemplo clásico de adaptación genética en respuesta a las presiones ambientales. En las islas Galápagos, los pinzones han cambiado varias formas de pico para explotar diferentes fuentes de alimento. Durante los periodos de sequía, cuando las semillas duras son la principal fuente de alimento, los pinzones con picos más grandes y fuertes tienen más probabilidades de tener una ventaja selectiva y reproducirse. Por el contrario, cuando el entorno se decanta por alimentos más blandos, los pinzones con picos más pequeños y ágiles tienen una ventaja selectiva. Estas adaptaciones son el resultado de cambios en genes específicos, como el gen aristaless-like homeobox 1 (ALX1), que influye en la forma del pico, y el gen high mobility group AT-hook 2 (HMGA2), que afecta al tamaño del pico.
Los cambios en el entorno actúan sobre estas variaciones genéticas, dando lugar a una diversidad de formas de pico adaptadas a diferentes nichos ecológicos. A lo largo de generaciones, estas adaptaciones genéticas permiten a los pinzones explotar eficazmente los recursos disponibles, lo que demuestra cómo los cambios genéticos pueden dar lugar a diversas formas y tamaños de pico en respuesta a los retos ambientales. Los pinzones viven en las islas Galápagos desde hace unos dos millones de años. A pesar de este largo periodo, han seguido siendo pinzones y no se han transformado en una especie diferente (es decir, no ha habido macroevolución).

Fig. 3.11. Picos de los pinzones de Galápagos
En conclusión, la ‘teoría de la evolución’ de Darwin debería llamarse ‘teoría de la adaptación genética’, ya que no hay pruebas convincentes de macroevolución. La microevolución se refiere a cambios a pequeña escala en las frecuencias alélicas dentro de una población a lo largo del tiempo, mientras que la adaptación genética describe específicamente cambios que mejoran la capacidad de un organismo para sobrevivir y reproducirse en su entorno. Por tanto, al hablar de cambios que confieren una ventaja de supervivencia, el término ‘adaptación genética’ sería más apropiado y preciso.
d. ¿Hemos evolucionado a partir de los simios?
Los antropólogos sugieren que la evolución humana comenzó a partir de los Hominoidea hace unos 20,4 millones de años. Los Hominoidea divergieron en Hominidae y Hylobatidae (gibones). A continuación, los Hominidae se dividieron en Homininae y Ponginae (orangutanes). Los Homininae se dividieron a su vez en Hominini y Gorillini (gorilas). Los Hominini se dividieron en Hominina (Australopithecina) y Panina (chimpancés). Los Hominina acabaron dividiéndose en Australopithecus y Ardipithecus. Los humanos evolucionaron a partir del Australopithecus hace unos 2,5 millones de años a través del Homo habilis, el Homo erectus y el Homo sapiens.

Fig. 3.12. ¿Hemos evolucionado a partir de los simios?
Discutamos si los humanos podrían haber evolucionado a partir de los Australopithecus (simios) mediante cambios genéticos en los últimos 2,5 millones de años. Existen mapas genéticos humanos, pero no se dispone de mapas genéticos de Australopithecus. Lucy, la Australopithecus más famosa, tenía un cerebro de tamaño comparable al de los chimpancés modernos. Por lo tanto, supongamos que los genes del Australopithecus son similares a los de los chimpancés. Las secuencias de ADN de los humanos y los chimpancés difieren en aproximadamente un 1,23% debido a los polimorfismos de nucleótido único (SNP), que son cambios de un solo par de bases en la secuencia de ADN. Si se tienen en cuenta las inserciones y deleciones (indels) de pares de bases en el genoma, la diferencia total aumenta. Los indels son segmentos de ADN que están presentes en una especie pero ausentes en la otra. Pueden representar un 3% más de diferencia en el genoma. En general, aunque los humanos y los chimpancés comparten alrededor del 98-99% de sus secuencias de ADN, el 1-2% de diferencia restante, junto con las variaciones en la regulación de los genes, explican las importantes diferencias físicas, cognitivas y de comportamiento entre las dos especies.
Se sabe que la tasa de mutación en los chimpancés es de aproximadamente 1 mutación por cada 100 millones de pares de bases por generación, comparable a la tasa de mutación en los humanos. Si suponemos que una generación de Australopithecus dura 25 años, habrán pasado 100.000 generaciones en 2,5 millones de años. Durante este periodo, la tasa de mutación total sería del 0,1% (100.000 / 100 millones). Esta tasa de mutación sólo representa el 10% de la diferencia genética entre humanos y chimpancés. Por tanto, parece poco probable que el Australopithecus pudiera evolucionar hasta convertirse en humanos en 2,5 millones de años. Esta estimación supone que todas las mutaciones son beneficiosas, aunque la mayoría de las mutaciones son perjudiciales.
Este argumento también puede examinarse considerando la alteración de codones mediante mutaciones genéticas aleatorias. Tanto los humanos como los chimpancés tienen aproximadamente entre 20.000 y 25.000 genes codificadores de proteínas. Debido al splicing alternativo y a las modificaciones postraduccionales, cada gen puede producir múltiples variantes proteicas, lo que da lugar a unas 80.000 a 100.000 proteínas funcionales únicas. El número de aminoácidos en las proteínas humanas oscila entre 20 y 33.000. Suponiendo que el 1% de los genes difieran entre humanos y chimpancés, y que ambas especies tengan 20.000 genes codificadores de proteínas con una media de 100 aminoácidos por proteína, cabría esperar que cada proteína de los chimpancés requiriera una mutación aminoacídica para igualarse a su homóloga humana.
Para que estas mutaciones se produjeran en el ADN del chimpancé, tendrían que evitar mutar los codones a codones de parada (UAA, UAG, UGA) entre los 64 codones posibles, ya que tales cambios darían lugar a proteínas no funcionales. La probabilidad de alcanzar esta tasa de mutación del 1% en 20.000 proteínas sin mutar en codones de parada ni en el propio codón del chimpancé es de (60/64)20000 = 10-561. Incluso sin tener en cuenta las mutaciones frameshift (inserciones o deleciones de nucleótidos), esta probabilidad es extraordinariamente baja y prácticamente imposible que se produzca por azar. Este argumento sugiere que los cambios macroevolutivos, como la transición del australopiteco al ser humano, son prácticamente imposibles a través de mutaciones aleatorias.
e. Diseño inteligente
El diseño inteligente, a menudo considerado sinónimo de creacionismo, es la teoría científica según la cual el universo y los organismos vivos se explican mejor por una causa inteligente que por procesos no dirigidos, como la selección natural o el azar. Un caso notable relacionado con el diseño inteligente es el juicio ante un tribunal federal celebrado en 2005 en Dover, Pensilvania (EE.UU.). Este juicio comenzó cuando unos padres presentaron una demanda alegando que la enseñanza del diseño inteligente en las escuelas públicas violaba la Constitución. Los padres argumentaban que el diseño inteligente es intrínsecamente religioso por naturaleza y que enseñarlo en las escuelas públicas contravenía la Cláusula de Establecimiento de la Constitución de EE.UU., que ordena la separación de Iglesia y Estado.
Durante el juicio, los partidarios del diseño inteligente y de la evolución presentaron sus respectivos argumentos. Una figura destacada que representaba el diseño inteligente era el bioquímico Michael Behe, de, quien afirmaba que las complejas estructuras de los organismos vivos no podían explicarse únicamente por la selección natural y sugería la posibilidad de que ciertas características hubieran sido moldeadas por una causa inteligente.
Sin embargo, el tribunal rechazó los argumentos de Behe y otros defensores del diseño inteligente, aceptando en su lugar las posturas de los defensores de la evolución. El juez dictaminó que la enseñanza del diseño inteligente era inconstitucional, declarando así ilegal la enseñanza del diseño inteligente en las escuelas públicas de Dover.
El principal problema de esta sentencia radica en la aceptación acrítica por parte del tribunal de los argumentos de los defensores de la evolución y los artículos científicos relacionados. Estos documentos suponen implícitamente que la vida surgió por azar y malinterpretan la adaptación genética al entorno como prueba de evolución. Sin embargo, como se resume en la Tabla 3.2, las teorías evolutivas sólo se aplican a los organismos vivos existentes y no pueden explicar el origen de la vida. Además, las teorías evolutivas se limitan a describir el comportamiento de genes que ya están integrados en el código genético. Sin embargo, el tribunal no tuvo en cuenta estos hechos científicos en su decisión, lo que plantea serias dudas sobre la imparcialidad del fallo.
William Paley, filósofo del siglo XVIII, es una figura fundacional de este argumento, que ilustró célebremente con su analogía del relojero. Paley sostenía que, al igual que la complejidad de un reloj implica un diseñador, la complejidad de la vida y del universo también implica un Creador divino. Sus ideas sentaron las bases de la moderna teoría del diseño inteligente. Los conceptos clave del diseño inteligente son la complejidad especificada, la complejidad irreducible y el ajuste fino. En los capítulos 1 y 2 se mostraron varios ejemplos de ajuste fino. Examinemos ahora en detalle la complejidad especificada y la complejidad irreducible.
i. Complejidad especificada
La complejidad específica, un concepto clave en el diseño inteligente, postula que ciertos patrones de la naturaleza son a la vez muy complejos y están ordenados específicamente para cumplir una función concreta, lo que indica un diseño intencionado. A diferencia de la complejidad aleatoria, la complejidad específica no sólo es intrincada, sino que también está ordenada de forma que consigue un resultado concreto. Esta doble característica sugiere que es improbable que tales patrones hayan surgido por casualidad.
Uno de los ejemplos de complejidad especificada es la estructura del ADN. La secuencia de nucleótidos en el ADN es muy compleja, con miles de millones de combinaciones potenciales incluso en una sola cadena. Esta complejidad garantiza que la disposición no es el resultado de procesos simples y aleatorios. Los mecanismos de replicación y reparación del ADN ponen aún más de relieve su complejidad. En estos procesos intervienen múltiples proteínas y enzimas que trabajan coordinadamente para copiar y mantener con precisión la información genética. La secuencia de nucleótidos no sólo es compleja, sino también muy específica, ya que codifica instrucciones precisas para sintetizar proteínas. Cada gen de la secuencia de ADN corresponde a una proteína concreta, e incluso pequeños cambios en la secuencia pueden afectar significativamente a la función de la proteína resultante. El ADN también contiene elementos reguladores que controlan cuándo y dónde se expresan los genes, lo que añade otra capa de especificidad a su función.
Es poco probable que la complejidad específica observada en el ADN haya surgido a través de procesos no dirigidos como las mutaciones aleatorias y la selección natural. En cambio, sugiere que una causa inteligente es una explicación más plausible para el origen de una información tan intrincada y funcionalmente específica.
Otro ejemplo de complejidad especificada es el flagelo bacteriano, una estructura motorizada en forma de látigo utilizada por ciertas bacterias para su locomoción. A continuación se explica por qué el flagelo bacteriano se considera un ejemplo de complejidad especificada.
El flagelo bacteriano está compuesto por unas 40 proteínas diferentes que forman diversos componentes, como el filamento, el gancho y el cuerpo basal. El cuerpo basal funciona como un motor rotativo, con un rotor, un estator, un eje de transmisión y una hélice. Para que el flagelo funcione, todas estas piezas deben estar presentes y correctamente ensambladas. La ausencia de cualquiera de estos componentes hace que el flagelo no funcione, lo que pone de manifiesto su complejidad.

Fig. 3.13. Flagelo bacteriano
Para que funcione, los componentes del flagelo deben estar dispuestos de una manera muy específica en. Las proteínas deben ensamblarse en una secuencia precisa y sus formas deben encajar exactamente, como las piezas de una máquina bien diseñada. El flagelo no sólo es complejo, sino que además cumple una función muy específica: propulsar a la bacteria. Funciona a velocidades extraordinarias, puede cambiar de dirección y consume poca energía, todo lo cual apunta a un diseño intencionado.
La complejidad específica del flagelo bacteriano no puede explicarse adecuadamente mediante mutaciones aleatorias y selección natural. La probabilidad de que un sistema tan integrado y funcional surja por casualidad es extremadamente baja. Además, dado que las formas intermedias del flagelo probablemente no serían funcionales, la vía evolutiva tradicional de mejoras graduales paso a paso parece poco plausible.
El flagelo también ejemplifica la complejidad irreducible, un subconjunto de la complejidad especificada, como se detallará en la siguiente sección. El argumento es que todas las partes del flagelo son necesarias para su función y, por lo tanto, no podría haber evolucionado mediante ligeras modificaciones sucesivas, como sugiere la evolución darwiniana.
ii. Complejidad irreductible
La complejidad irreductible es un concepto introducido por el bioquímico Michael Behe, que postula que ciertos sistemas biológicos son demasiado complejos para haber evolucionado mediante modificaciones graduales, paso a paso. Estos sistemas, como el flagelo bacteriano o la cascada de coagulación de la sangre, constan de múltiples partes interdependientes que deben estar presentes y funcionar para que el sistema funcione. Si se elimina una de ellas, el sistema deja de funcionar. Estas estructuras intrincadas e interdependientes de indican la presencia de un diseñador inteligente, ya que no pueden explicarse únicamente por la selección natural y la mutación aleatoria. Este concepto desafía la teoría evolutiva convencional y apoya la idea de un diseño intencionado en la naturaleza.
Un ejemplo de complejidad irreducible es el ciclo visual, un proceso bioquímico en el ojo que convierte la luz en señales eléctricas, permitiendo la visión. Este sistema consta de múltiples partes interdependientes que deben estar todas presentes y funcionar para que el proceso sea eficaz. Si faltara algún componente o no funcionara, todo el ciclo visual fallaría, lo que ilustra el concepto de complejidad irreducible. Los componentes clave del ciclo visual son los fotorreceptores (bastones y conos), la rodopsina, las opsinas, la retina, la vía de transducción de señales y el procesamiento neural.

Fig. 3.14. Etapas moleculares del ciclo visual
Los fotorreceptores son células de la retina que detectan la luz. Los bastones son responsables de la visión con poca luz, mientras que los conos detectan el color. Cada fotorreceptor contiene moléculas sensibles a la luz llamadas fotopigmentos, principalmente rodopsina en los bastones. Este fotopigmento en los bastones está formado por una proteína llamada opsina y una molécula sensible a la luz llamada retinal. Los conos contienen diferentes opsinas que responden a varias longitudes de onda de la luz, lo que permite la visión en color. El retinal, un derivado de la vitamina A, cambia de forma cuando absorbe la luz. Este cambio de forma activa la opsina, iniciando la cascada de transducción visual. La opsina activada activa a su vez una proteína G llamada transducina. La transducina activa la fosfodiesterasa (PDE), que disminuye el nivel de GMP cíclico (cGMP) en la célula. La disminución del GMPc cierra los canales iónicos de la membrana de la célula fotorreceptora, lo que provoca la hiperpolarización de la célula y genera una señal eléctrica. La señal eléctrica se transmite a través de las células bipolares a las células ganglionares, que envían la señal al cerebro a través del nervio óptico. El cerebro procesa estas señales para formar imágenes visuales.
Cada componente del ciclo visual es interdependiente. Los fotorreceptores, la rodopsina, la retina, la transducina, la PDE y los canales iónicos deben estar presentes y funcionar correctamente para que se produzca la visión. La eliminación de cualquiera de estos componentes provocaría el fallo del sistema. Podemos argumentar que un sistema tan complejo no podría haber evolucionado a través de una serie de pequeños cambios incrementales porque las etapas intermedias sin todos los componentes no serían funcionales y, por tanto, no se verían favorecidas por la selección natural. Las intrincadas vías bioquímicas y las precisas interacciones moleculares que intervienen en el ciclo visual ponen de manifiesto la complejidad y especificidad necesarias para la visión. La naturaleza interdependiente de sus componentes y la complejidad de los procesos bioquímicos implicados sugieren que este sistema no pudo surgir mediante procesos evolutivos no dirigidos, sino que apunta a un diseñador inteligente, el Creador divino.
El ciclo visual en términos de un programa informático puede ayudar a ilustrar su complejidad y sus procesos interdependientes. He aquí una analogía conceptual utilizando python:
Ciclo visual escrito en programa informático
# inicialización: configura el entorno para el ciclo visual, incluidos los fotorreceptores (bastones y conos)
clase CicloVisual:
def __init__(self):
self.fotorreceptores = {'bastones': [], 'conos': []}
self.inicializar_fotopigmentos()
self.ruta_señal_activa = False
# entrada de usuario: detecta la luz entrante e inicia el proceso de activación de los fotopigmentos
def detect_light(self, light_wavelength):
if light_wavelength in visible_spectrum:
self.activate_photopigment(light_wavelength)
# evento desencadenante: cambia la forma de la retina y activa la opsin, que a su vez activa la vía de transducción de señales
def activate_photopigment(self, wavelength):
retinal = self.change_retinal_shape(wavelength)
opsina = self.bind_retinal_to_opsin(retinal)
self.start_signal_transduction(opsin)
# gestión de eventos: activa la transducina y la PDE, lo que provoca una reducción de los niveles de GMPc, el cierre de los canales iónicos y la generación de una señal eléctrica
def start_signal_transduction(self, opsin):
self.signal_pathway_active = True
transducina = self.activate_transducin(opsina)
PDE = self.activate_pde(transducina)
self.regulate_cGMP_levels(pde)
self.generate_electrical_signal()
# manejo de señales: ajusta los canales iónicos en función de los niveles de GMPc para facilitar la generación de señales eléctricas
def regulate_cGMP_levels(self, pde):
cGMP_level = self.reduce_cGMP(pde)
self.adjust_ion_channels(cGMP_level)
# señal de salida: crea y transmite la señal eléctrica al cerebro
def generate_electrical_signal(self):
if self.signal_pathway_active:
electrical_signal = self.create_signal()
self.transmit_signal_to_brain(electrical_signal)
# comunicación en red: procesa y reenvía la señal a través de las células bipolares y ganglionares, enviándola finalmente a través del nervio óptico
def transmit_signal_to_brain(self, signal):
bipolar_cells = self.process_signal_with_bipolar_cells(signal)
ganglion_cells = self.forward_signal_to_ganglion(bipolar_cells)
optic_nerve = self.send_signal_via_optic_nerve(ganglion_cells)
self.visual_perception(optic_nerve)
# resultado final: el cerebro decodifica y procesa la señal para crear una imagen visual
def visual_perception(self, optic_nerve):
corteza_visual = self.decodificar_señal(nervio_óptico)
self.render_imagen(corteza_visual)
Esta analogía ilustra los pasos interdependientes y la complejidad del ciclo visual, muy parecido a un programa informático con varias funciones y controladores de eventos que trabajan juntos para lograr un resultado específico. Si omitimos alguno de los pasos o los utilizamos en el orden incorrecto, no obtendremos el resultado deseado. El hecho de que el ciclo visual pueda representarse en como un programa informático sugiere que el ojo fue diseñado de forma inteligente. El proyecto de diseño del ojo está vinculado al gen PAX6, situado en el cromosoma 11, que desempeña un papel crucial en el desarrollo del ojo.
iii. Libros notables sobre el diseño inteligente
Evolution: A Theory in Crisis (Michael Denton: 1985): Denton critica la evolución darwiniana, argumentando que la complejidad de los sistemas biológicos no puede explicarse adecuadamente sólo mediante la selección natural. Denton presenta pruebas procedentes de diversos campos, como la biología molecular y la paleontología, para poner de relieve las lagunas e incoherencias de la teoría evolutiva. Sostiene que las intrincadas estructuras y funciones observadas en los organismos vivos apuntan a un diseño inteligente y no a mutaciones y selección aleatorias. El libro desafía el consenso científico imperante y sugiere que se necesita una explicación alternativa para explicar el origen y la diversidad de la vida.
La caja negra de Darwin: El desafío bioquímico a la evolución (Michael J. Behe: 2006): En este libro seminal, Michael Behe introduce el concepto de complejidad irreducible, argumentando que ciertos sistemas biológicos, como el flagelo bacteriano, son demasiado complejos para haber evolucionado únicamente a través de la selección natural. Behe sostiene que estos sistemas se explican mejor mediante el diseño inteligente. El libro cuestiona la idoneidad de la evolución darwiniana para explicar la intrincada maquinaria de la vida a nivel molecular y ha suscitado un importante debate tanto en círculos científicos como filosóficos.
Darwin a prueba (Phillip Johnson: 2010): Este libro critica los fundamentos científicos de la evolución darwiniana. Johnson, profesor de Derecho, examina las pruebas de la evolución con el escrutinio de un analista jurídico. Sostiene que la selección natural y la mutación aleatoria no explican adecuadamente la complejidad de la vida. Johnson sugiere que gran parte del apoyo al darwinismo se basa en el naturalismo filosófico más que en la ciencia empírica. Cuestiona la reticencia de la comunidad científica a considerar explicaciones alternativas, como el diseño inteligente, y aboga por un debate más abierto sobre los orígenes de la vida. El libro influye en la promoción del diseño inteligente y cuestiona el predominio de la teoría darwiniana en biología.
Signature in the Cell : DNA and the Evidence for Intelligent Design (Stephen C. Meyer, 2010): Este libro explora los orígenes de la vida y la información codificada en el ADN. Meyer sostiene que la información compleja y específica del ADN se explica mejor por una causa inteligente, ya que los procesos naturalistas no logran explicar el origen de dicha información. Presenta argumentos detallados a favor del diseño inteligente basándose en las complejidades de la información genética, y sugiere que el origen de la vida apunta a una creación intencionada y no a procesos aleatorios.
Darwin evoluciona : The New Science About DNA That Challenges Evolution (Michael J. Behe, 2020): Otro libro de Behe sostiene que los recientes descubrimientos genéticos socavan la evolución darwiniana tradicional. Afirma que, aunque la selección natural y las mutaciones aleatorias pueden explicar adaptaciones menores, no dan cuenta de la complejidad de la maquinaria molecular dentro de las células. Introduce el concepto de ‘involución’, según el cual las mutaciones conducen a la pérdida de información genética en lugar de a la creación de nuevos rasgos beneficiosos. Behe sostiene que estas limitaciones genéticas apuntan a la necesidad de un diseñador inteligente, cuestionando el marco evolutivo tradicional y proponiendo que el diseño inteligente ofrece una explicación más plausible de la complejidad de la vida.
El misterio del origen de la vida: Reassessing Current Theories (Charles B. Thaxton et al., 2020): Este innovador trabajo critica las diversas teorías naturalistas del origen de la vida y propone el diseño inteligente como explicación más plausible. Argumentan que la química prebiótica y la formación de la vida a partir de la no vida se explican mejor por una causa inteligente. El libro analiza las deficiencias de las teorías contemporáneas sobre el origen de la vida y presenta el diseño inteligente como una alternativa científicamente viable, sentando las bases del moderno movimiento del diseño inteligente.
La inferencia de diseño : Eliminating Chance through Small Probabilities (William A. Dembski y Winston Ewert, 2023): Este libro sienta las bases teóricas para detectar el diseño en la naturaleza. Exploran el marco matemático para detectar el diseño inteligente. Los autores presentan el argumento de que los sistemas complejos que muestran una complejidad especificada se explican mejor por una causa inteligente que por procesos aleatorios. Introducen el concepto de ‘complejidad especificada’, que combina la complejidad con un patrón dado de forma independiente. El libro utiliza la teoría de la probabilidad para demostrar que ciertos patrones de la naturaleza son demasiado improbables para haber surgido por azar. Mediante un análisis riguroso, Dembski y Ewert sostienen que reconocer el diseño es una práctica científica legítima y proporciona herramientas para distinguir el diseño del azar en los sistemas biológicos.
f. Física de partículas y creación
En la sección anterior, exploramos el origen de la vida analizando sus componentes fundamentales, como los aminoácidos, el ARN, las proteínas, el ADN y las células. Estos componentes están formados por átomos, que implícitamente suponemos que existen de forma natural. Los átomos están formados por partículas elementales. En esta sección analizaremos más detenidamente el origen de estas partículas, estudiando si surgieron espontáneamente o se formaron mediante un proceso intencionado.
Según el Modelo Estándar de la física de partículas, toda la materia del universo está compuesta por 17 partículas elementales. Se trata de 6 quarks, 6 leptones, 4 bosones gauge (gluones, fotones, bosones Z y bosones W) y el bosón de Higgs. Cada una de estas partículas tiene propiedades específicas, como la masa, la carga y el espín, y cada una desempeña un papel único en las interacciones entre partículas, de forma similar a como los orgánulos de una célula desempeñan funciones distintas.

Fig. 3.15. Las partículas elementales del Modelo Estándar
Los quarks son componentes fundamentales de la materia, esenciales en la formación de protones y neutrones. Los protones están formados por dos quarks ‘up’ y un quark ‘down’, mientras que los neutrones están formados por un quark ‘up’ y dos quarks ‘down’. Los quarks se mantienen unidos por la fuerza fuerte, mediada por los gluones. A diferencia de las fuerzas gravitatoria o electromagnética, que disminuyen con la distancia, la fuerza fuerte entre quarks aumenta a medida que se alejan y disminuye a medida que se acercan, manteniendo una separación específica. Los quarks pueden cambiar de tipo durante las interacciones de partículas, como la desintegración beta, en la que un neutrón se transforma en protón al convertir un quark down en un quark up.
Los bosones gauge son partículas fundamentales que median las fuerzas básicas de la naturaleza. Entre ellas se encuentran el fotón para la fuerza electromagnética, los bosones W y Z para la fuerza débil y el gluón para la fuerza fuerte. Cada bosón gauge está asociado a un campo específico y transmite la fuerza entre partículas. Son esenciales para explicar las interacciones a nivel cuántico, ya que rigen la forma en que las partículas interactúan y se unen para formar la materia.
El mecanismo de Higgs es un proceso que explica cómo las partículas elementales adquieren masa. En él interviene el campo de Higgs, un campo de energía que impregna el universo. Cuando las partículas interactúan con el campo de Higgs, adquieren masa, de forma similar a como los objetos que se mueven a través de un medio experimentan resistencia. El bosón de Higgs, una partícula asociada al campo de Higgs, fue descubierto en 2012, confirmando esta teoría. Sin el mecanismo de Higgs, las partículas seguirían sin masa y el universo carecería de la estructura necesaria para la formación de átomos, organismos vivos, planetas y estrellas.
La física de partículas opera a un nivel increíblemente avanzado e intrincado, ofreciendo profundos conocimientos sobre la naturaleza y los orígenes del universo. Esto nos lleva a plantearnos, entre otras muchas, las siguientes preguntas fundamentales:
- ¿Cómo se crearon las 17 partículas fundamentales con propiedades tan precisas?
- ¿Cómo adquirieron los bosones gauge la propiedad de mediación de fuerzas?
- ¿Cómo se originó el mecanismo de Higgs?
- ¿Cómo se originó el mecanismo de desintegración beta?
- ¿Cómo se pueden describir matemáticamente las propiedades de las partículas elementales?
Si las respuestas a las preguntas anteriores fueran puramente el resultado de procesos aleatorios, el mundo tal y como lo conocemos podría no existir. Por ejemplo, si faltara una sola partícula fundamental, si no se hubiera establecido el mecanismo de Higgs o si los valores de masa y espín de las partículas elementales fueran ligeramente diferentes, los neutrones, protones y electrones no podrían mantenerse unidos. El resultado sería el colapso de toda la materia, lo que haría imposible la formación de cualquier cosa, incluidos los seres humanos. Esta precisión en la estructura fundamental del universo ejemplifica el concepto de ‘complejidad irreducible’ en el ámbito de la física de partículas, un principio que a menudo se asocia con el diseño inteligente.
La creación de partículas elementales para formar materia puede compararse a la formación de células y orgánulos en los organismos multicelulares. Al igual que las células y los orgánulos específicos tienen funciones y propiedades distintas que contribuyen a la compleja funcionalidad de los seres vivos, las partículas elementales poseen características precisas que permiten la formación de átomos, moléculas y, en última instancia, todas las materias. Este paralelismo subraya la sofisticación y la intencionalidad inherentes al mundo natural, ya sea en el nivel microscópico de las células vivas, en el reino subatómico de las partículas fundamentales o en la escala macroscópica de los organismos vivos, las estrellas y las galaxias.
El hecho de que la formación de partículas elementales y sus interacciones puedan describirse con precisión utilizando las ecuaciones matemáticas de la mecánica cuántica sugiere que son el resultado de un diseño matemático intencionado y no de la mera casualidad. De lo contrario, tendríamos que suponer que las partículas elementales poseen inteligencia y la capacidad de determinar, por sí solas, los valores exactos de masa, carga y espín necesarios para formar materia e interactuar con otras partículas. Sin embargo, sabemos que no es así, ya que las partículas elementales no tienen conciencia ni una comprensión intrínseca de la mecánica cuántica.
El intrincado diseño y la coordinación que se observan tanto en los sistemas biológicos como en la física de partículas sugieren claramente la presencia de una inteligencia subyacente y de una creación intencionada (sello distintivo del diseño inteligente), en lugar de una serie de sucesos aleatorios.
g. Existencia de extraterrestres y creación
La posibilidad de que existan alienígenas, o vida extraterrestre, lleva décadas fascinando a científicos y público en general en.Dada la inmensidad del universo, con miles de millones de galaxias cada una de las cuales contiene miles de millones de estrellas y potencialmente aún más planetas, parece estadísticamente plausible que pudiera existir vida en otros lugares si ésta surgiera espontáneamente. El número de civilizaciones extraterrestres en una galaxia puede estimarse mediante Ecuación de Drake: N = R* × fp× ne× fl× fi× fc× L donde, N es el número de civilizaciones avanzadas, R* es la tasa de formación estelar, fp es la fracción de tener planetas, ne es el número de planetas que soportan vida, fl es la fracción de planetas donde se desarrolla la vida, fi es la fracción de planetas donde evoluciona la vida inteligente, fc es la fracción de civilizaciones que pueden enviar señales y L es el tiempo que las civilizaciones pueden comunicarse. Con un valor adecuado para cada parámetro, el número estimado de civilizaciones en una galaxia es aproximadamente 2.

Fig. 3.16. ¿Existen los extraterrestres?
Los proyectos de búsqueda de inteligencia extraterrestre (SETI) se iniciaron en 1960. Estos proyectos utilizan diversos métodos y tecnologías para escanear el cosmos en busca de pruebas de civilizaciones extraterrestres. Éstos son algunos de los principales proyectos SETI.
El Proyecto Ozma fue el primer experimento SETI moderno. Utilizó un radiotelescopio para escanear las estrellas Tau Ceti y Epsilon Eridani en busca de posibles señales extraterrestres. SETI@home era un proyecto informático distribuido que utilizaba la potencia de procesamiento ociosa de los ordenadores domésticos. Los voluntarios instalaban software en sus ordenadores personales para analizar las señales de radio en busca de señales de inteligencia extraterrestre. El Allen Telescope Array es una red de radiotelescopios diseñada para la búsqueda continua y sistemática de señales extraterrestres. Consta de varias pequeñas antenas parabólicas que trabajan conjuntamente para explorar amplias zonas del cielo. Breakthrough Listen es el proyecto SETI más completo hasta la fecha, cuyo objetivo es explorar un millón de las estrellas más cercanas y 100 galaxias cercanas en busca de posibles señales. El proyecto Fast Radio Burst investiga las misteriosas ráfagas de radio rápidas detectadas desde el espacio, que podrían aportar información sobre fenómenos cósmicos desconocidos. Láser SETI es un proyecto centrado en la detección de señales ópticas procedentes de civilizaciones extraterrestres, que explora la posibilidad de comunicación interestelar a través de transmisiones láser.
A pesar de las continuas búsquedas con radiotelescopios y telescopios ópticos avanzados, los proyectos SETI no lograron encontrar pruebas definitivas de vida extraterrestre inteligente.

Fig. 3.17. Radiotelescopios utilizados para SETI
Si existen numerosas civilizaciones extraterrestres, podrían habernos visitado o estar visitándonos ahora. En tal caso, ¿qué métodos de viaje espacial utilizarían? Viajar al espacio utilizando objetos voladores (cohetes u ovnis) se enfrenta a retos insuperables debido al enorme tamaño del universo. Incluso la estrella más cercana, Próxima Centauri, se encuentra a 4,24 años luz, por lo que se necesitarían decenas de miles de años para llegar a ella con la tecnología actual. Las enormes distancias hacen imposible explorar nuestra galaxia, por no hablar del universo, durante la vida humana.
Los posibles métodos avanzados de propulsión podrían incluir motores warp o viajes a través de agujeros de gusano. El motor warp es un concepto teórico de viaje espacial más rápido que la luz, inspirado en la relatividad general de Einstein. Propuesto por el físico Miguel Alcubierre en 1994, el motor warp consiste en crear una ‘burbuja warp’ que contrae el espacio delante de una nave espacial y expande el espacio detrás de ella. Esto permitiría a la nave moverse más rápido que la luz en relación con observadores externos sin violar las leyes de la física. El principal problema es que se necesita materia exótica con densidad de energía negativa, que aún no se ha descubierto ni creado. Aunque prometedor en teoría, se necesitan avances científicos y tecnológicos significativos para que un motor warp sea factible para su uso práctico en la exploración especial.
El viaje espacial a través de agujeros de gusano es un concepto teórico que implica atajos a través del espacio-tiempo que conectan puntos distantes en el universo. Predichos por la relatividad general de Einstein, los agujeros de gusano o puentes de Einstein-Rosen podrían permitir viajes instantáneos a través de enormes distancias cósmicas. Para su uso práctico, un agujero de gusano transitable tendría que estar estabilizado, lo que teóricamente requeriría materia exótica con densidad de energía negativa para evitar el colapso. A pesar de ser un tropo popular de la ciencia ficción, los agujeros de gusano siguen siendo especulativos, sin pruebas experimentales. Si fueran factibles, podrían revolucionar los viajes espaciales, permitiendo la exploración de galaxias lejanas y reduciendo el tiempo de viaje de años a meros instantes. Sin embargo, se necesitan importantes avances científicos y tecnológicos para hacer realidad este concepto.

Fig. 3.18. Agujero de gusano
El teletransporte a través del hiperespacio o la gran masa podría ser otro método para lograr el viaje instantáneo a través de grandes distancias eludiendo el espacio tridimensional convencional. El hiperespacio se refiere a una dimensión o serie de dimensiones adicionales que van más allá de las tres dimensiones espaciales y una dimensión temporal conocidas, proporcionando un atajo a través del tejido del universo. Del mismo modo, bulk es un término utilizado en teorías como la cosmología de branas dentro de la teoría de cuerdas, en la que nuestro universo se concibe como una ‘brana’ dentro de un espacio de dimensiones superiores llamado bulk. En estas teorías, el teletransporte consiste en desplazarse a través de estas dimensiones superiores para reaparecer instantáneamente en otro lugar de nuestro universo. Marcos teóricos como el modelo Randall-Sundrum proponen la existencia de esas dimensiones superiores que podrían permitir atajos a través del espacio-tiempo. Si tales dimensiones existen y se puede acceder a ellas, sería posible explotarlas para el teletransporte, evitando las limitaciones del viaje relativista y haciendo potencialmente factible el viaje más rápido que la luz.

Fig. 3.19. Teletransporte
Si la vida surge espontáneamente, como suponía la ecuación de Drake, el número total de civilizaciones extraterrestres en el universo sería de unos 400.000 millones (2 civilizaciones en cada una de 200.000 millones de galaxias). La vida en la Tierra comenzó hace aproximadamente 4.000 millones de años. Ahora, imaginemos que el 1% de las civilizaciones extraterrestres empezaron 1 millón de años antes que la nuestra y siguieron un camino evolutivo similar. En ese caso, su civilización sería 1 millón de años más avanzada que la nuestra. Con una ventaja tan significativa, podrían haber desarrollado tecnologías avanzadas de teletransporte, lo que les permitiría viajar a cualquier parte del universo con la misma facilidad con la que nosotros visitamos a nuestros vecinos. Si la población de una de estas civilizaciones es de 1.000 millones, el número total de alienígenas sería de un quintillón (1018). Si sólo el 1% de ellos pudiera visitar la Tierra durante un solo día cada 10 años, la Tierra estaría abarrotada de unos 10 billones de alienígenas cada día: 1.000 veces la población humana actual. Sin embargo, no hemos observado ninguna prueba de su presencia. ¿Cómo explicar esta aparente contradicción?
Este problema se conoce como la Paradoja de Fermi, llamada así en honor a Enrico Fermi, que se hizo famoso preguntando: ‘¿Dónde está todo el mundo?’. Las respuestas podrían ser: (i) la hipótesis (evolución) de la Ecuación de Drake es errónea, o (ii) civilizaciones avanzadas podrían utilizar tecnología indetectable con nuestros métodos actuales o evitar deliberadamente la detección. Si los extraterrestres no fueran ni bacterias ni seres invisibles, probablemente su existencia ya nos habría sido revelada de alguna manera. Sin embargo, el hecho de que aún no hayamos detectado ninguna prueba de su existencia sugiere que la hipótesis evolutiva de la Ecuación de Drake es muy probablemente incorrecta.
h. Instintos en los organismos vivos y en la creación
Los ordenadores se componen de tres elementos principales: hardware, software y firmware. El firmware es un software especializado programado en la ROM o UEFI, que proporciona un control crítico para un hardware específico y actúa como intermediario entre el hardware y el software. Es crucial para arrancar el sistema, gestionar las operaciones de hardware y garantizar la funcionalidad del dispositivo.
El firmware de los ordenadores y el instinto de los organismos vivos comparten una similitud clave: ambos son sistemas intrínsecos, preprogramados, que rigen funciones esenciales. El firmware inicializa y gestiona las operaciones, garantizando el correcto funcionamiento desde el encendido. Del mismo modo, el instinto es un patrón de comportamiento natural e innato que dirige las actividades de supervivencia, como alimentarse, aparearse o huir del peligro. Ambos sistemas funcionan automáticamente sin intervención consciente, proporcionando una guía fundamental para el funcionamiento eficaz y la respuesta al entorno.
En esencia, el firmware es para los ordenadores lo que el instinto es para los organismos vivos: un sistema preconfigurado e integrado en, esencial para el funcionamiento básico y la supervivencia. Al igual que los diseñadores de ordenadores incorporan el firmware en la memoria ROM, el Creador divino incorpora el instinto en el cerebro y el sistema nervioso de los organismos vivos. Permítanme mostrarles algunos ejemplos de instintos que ilustran este concepto.
i. Construcción de nidos de abejas albañiles
En el libro de Jean-Henri Fabre ‘Las abejas albañiles’ (parte del ‘Libro de los insectos’), describe el intrincado proceso de construcción de nidos de las abejas albañiles. Estas abejas seleccionan una superficie plana adecuada, a menudo una piedra, para iniciar su construcción. Recogen barro y pequeños guijarros, creando meticulosamente celdas para sus crías. La abeja hembra transporta gránulos de barro hasta el lugar, dándoles forma y compactándolos para formar una pared celular segura. A continuación, recoge néctar y polen para abastecer cada celda y pone un solo huevo antes de sellarla con más barro. Este proceso se repite, dando lugar a una serie de celdas de barro reforzadas con guijarros y ordenadamente dispuestas que protegen a las larvas en desarrollo. Las observaciones de Fabre ponen de relieve la notable precisión y diligencia de estas abejas solitarias.
Describe un experimento en el que cambió un nido inacabado por otro terminado. La abeja albañil, al volver y ver que su nido inacabado había sido sustituido por otro terminado, mostró un comportamiento interesante. En lugar de reanudar el trabajo en el nuevo nido, la abeja continuó su construcción como si no hubiera habido ningún cambio. No reconoció el nido terminado como obra suya y persistió en sus acciones habituales, trayendo barro y continuando la construcción.

Fig. 3.20. Abeja albañil construye nido encima ya terminado
Este experimento ilustra la naturaleza instintiva y programada del comportamiento de la abeja, impulsado por una secuencia interna de acciones más que por señales visuales del estado del nido.
Fabre realizó el experimento inverso cambiando un nido de abeja albañil terminado por otro inacabado. Observó que cuando la abeja albañil regresaba al lugar y encontraba el nido terminado sustituido por otro inacabado, no seguía trabajando en el nuevo nido incompleto. En cambio, la abeja parecía confundida y pasó un tiempo inspeccionando el nido alterado, pero finalmente no reanudó la construcción. Entonces pasa a la siguiente acción de llenarlo de miel, aunque esté rebosante. Este comportamiento demuestra el fuerte apego de la abeja albañil a su nido específico y la dificultad de adaptarse a cambios inesperados en su entorno. Este experimento también pone de relieve la naturaleza instintiva del proceso de construcción de nidos de la abeja albañil.

Fig. 3.21. Abeja albañil llena de miel un nido inacabado
Fabre hizo otro experimento interesante. La abeja albañil llena primero su nido de néctar y, a continuación, gira 180 grados y se quita el polen de las patas y el cuerpo. Si la interrumpen cuando está a punto de desempolvar el polen, vuela y espera a que pase la amenaza. Cuando regresa al nido, vuelve a empezar desde el principio. Llena su nido de néctar aunque no haya nada en su saco de néctar. Este experimento demuestra que las abejas siguen instintivamente un programa incorporado de recolección de néctar, y su secuencia de acciones no puede alterarse.

Fig. 3.22. Comportamiento de la abeja albañil cuando es perturbada
Cuando la abeja albañil termina de construir su nido, lo llena de néctar y polen, pone su huevo en él y luego sella la parte superior del nido. La parte superior sellada es tan dura como el cemento, Fabre realizó otro experimento: para un nido, pegó papel en la parte superior, y para otro, colocó un cono de papel encima. Observó el comportamiento de las abejas nacidas. En el nido con papel pegado, la abeja utilizó sus fuertes mandíbulas para cortar la parte superior sin ningún problema. En el nido con un cono de papel, cortó la parte superior pero no sabía qué hacer a continuación. Esperaba ver el cielo abierto, pero el cono de papel la desorientó, no intentó perforarlo y finalmente murió.

Fig. 3.23. Nido pegado con papel y cubierto con un cono de papel
Todos los experimentos anteriores demuestran la naturaleza instintiva y programada del comportamiento de la abeja albañil, impulsado por una secuencia interna de acciones incrustadas en su código genético.
ii. Construcción de nidos de aves tejedoras
El ave tejedor, conocido por sus intrincados y elaborados nidos, teje hábilmente briznas de hierba y otros materiales vegetales en complejas estructuras, haciendo gala de una notable artesanía e ingeniería instintiva.

Fig. 3.24. Nido de ave tejedor
Eugène Marais, naturalista y poeta sudafricano, realizó fascinantes experimentos con aves tejedoras para estudiar su comportamiento en la construcción de nidos y el papel del instinto. Marais pretendía averiguar si las intrincadas habilidades de construcción de nidos de los aves tejedores eran puramente instintivas o si implicaban un comportamiento aprendido.
Marais crió a las tejedoras aisladas de su entorno natural para que no estuvieran expuestas a otras aves ni a actividades de construcción de nidos. Observó a estas aves aisladas desde la eclosión hasta la madurez, asegurándose de que no tuvieran la oportunidad de aprender de otras aves tejedoras durante cuatro generaciones. Para la quinta generación, Marais les proporcionó los mismos materiales que las aves tejedoras salvajes utilizan para construir sus nidos, como hierba y ramitas. A pesar de no haber visto nunca un nido ni a otros aves construyéndolo, los tejedores aislados empezaron a construir nidos casi idénticos a los de sus congéneres salvajes. Mostraron las mismas técnicas intrincadas de tejido, métodos de anudado y estructura general. Los nidos construidos por estas aves aisladas mostraban características de diseño típicas de su especie, lo que indica que sus habilidades para construir nidos eran innatas y no aprendidas por observación o mimetismo.
Marais llegó a la conclusión de que el complejo comportamiento de construcción de nidos de las aves tejedoras está impulsado por el instinto. Este comportamiento innato está codificado en su cerebro y sistema nervioso, lo que les permite construir nidos elaborados sin experiencia ni aprendizaje previos. Estos comportamientos innatos se diseñan a propósito y se transmiten de generación en generación a través del ADN.
iii. Formación de la concha del Nautilus
El nautilo es un molusco marino conocido por su bella y característica concha. La forma de su concha sigue una precisa espiral logarítmica. La formación de la concha del nautilo es otro ejemplo notable de instinto, que implica una compleja interacción de procesos biológicos y químicos intrincadamente coordinados para producir su estructura única.
El proceso comienza cuando el nautilo es todavía un embrión dentro de un huevo. Durante esta etapa se forma la concha inicial, denominada protoconcha. Esta primera cámara es pequeña y sirve de base para el posterior crecimiento de la concha. El manto, un tejido especializado que recubre la concha, segrega capas de carbonato cálcico (CaCO3) en forma de aragonito, una estructura cristalina. Las células del manto extraen iones de calcio del agua de mar y los combinan con iones de carbonato para formar carbonato cálcico. El manto también segrega una matriz orgánica compuesta de proteínas y polisacáridos, que sirve de andamiaje para la deposición del carbonato cálcico. Esta matriz ayuda a controlar la forma y orientación de los cristales de aragonito, garantizando la resistencia y durabilidad de la concha.

Fig. 3.25. Concha de nautilus que muestra un patrón espiral logarítmico.
A medida que crece, el nautilo añade periódicamente nuevas cámaras a su concha. Cada nueva cámara es más grande que la anterior, para acomodar el creciente tamaño del nautilo. El nautilo avanza en la concha y cierra las cámaras más antiguas con una pared llamada tabique, creando una serie de cámaras interconectadas cada vez más grandes. Un órgano especializado llamado sifúnculo recorre todas las cámaras de la concha. Esta estructura tubular regula el contenido de gas y líquido de las cámaras. Al regular los niveles de gas (principalmente nitrógeno) y líquido, el sifúnculo ayuda al nautilo a controlar su flotabilidad, lo que le permite moverse hacia arriba y hacia abajo en la columna de agua. La capa más externa de la concha, conocida como periostraco, es una capa orgánica que protege las capas subyacentes de carbonato cálcico de la disolución y los daños físicos. Debajo del periostracum hay capas de aragonito, dispuestas en una estructura nacarada o prismática, que contribuyen a la iridiscencia y resistencia de la concha.
La intrincada coordinación necesaria para la secreción de carbonato cálcico, la formación de cámaras y la regulación de la flotabilidad a través del sifúnculo indica un sistema ‘todo o nada’ demasiado complejo para haber surgido por evolución gradual. La ausencia de fósiles transicionales claros en el registro, unida a la etiqueta de ‘fósil viviente’ del nautilus, implica una aparición repentina y sugiere que la sofisticada formación de su concha apunta a una creación intencionada más que a una evolución no dirigida. El nautilus no posee conocimientos matemáticos ni bioquímicos; por lo tanto, la formación precisa de la forma logarítmica de su concha, la compleja regulación bioquímica de la secreción de la concha y la perfecta integración de su sistema de flotación no son el resultado de procesos aleatorios. Por el contrario, estas características sugieren la existencia de un plan genético preprogramado que permite al nautilo construir su intrincada concha con notable precisión, lo que refuerza la idea de un diseño intencionado y no de una evolución no guiada.
i. Las matemáticas en la naturaleza y la creación
‘Las matemáticas son el lenguaje en el que Dios ha escrito el universo’. - Galileo Galilei
En la naturaleza abundan los patrones y principios matemáticos, como la proporción áurea, el ángulo de oro, la secuencia de Fibonacci, la espiral logarítmica y los fractales.

Fig. 3.26. Proporción áurea, ángulo áureo, espiral logarítmica y fractal
- La proporción áurea, a menudo denotada por la letra griega φ (= (a+b)/a=a/b), es un número irracional aproximadamente igual a 1,618. Se produce cuando el cociente entre dos cantidades es igual al cociente entre su suma y la mayor de las dos cantidades.
- El ángulo áureo es el ángulo subtendido por dos radios que dividen un círculo en dos longitudes de arco en la proporción áurea. Es el menor de los dos ángulos (~137,5 grados) creados al dividir la circunferencia de un círculo según la proporción áurea
- La sucesión de Fibonacci es una serie de números en la que cada número es la suma de los dos anteriores, empezando por 0 ó 1 (por ejemplo, 0, 1, 1, 2, 3, 5, 8, ...).
- La espiral logarítmica es una curva en espiral autosimilar que aparece con frecuencia en la naturaleza. Se caracteriza por la propiedad de que el ángulo entre la tangente y la línea radial en cualquier punto es constante.
- Los fractales son patrones complejos autosemejantes a diferentes escalas. Suelen crearse repitiendo un proceso simple una y otra vez en un bucle de retroalimentación continua.
Exploremos dónde se encuentran estos principios matemáticos en la naturaleza. La filotaxis es la disposición de hojas, flores u otras estructuras botánicas en el tallo de una planta. Es un concepto clave en botánica y refleja la forma en que las plantas maximizan su exposición a la luz solar y otros recursos ambientales. La disposición de las hojas sigue la secuencia de Fibonacci, donde el número de hojas en espirales sucesivas es un número de Fibonacci. Los posibles patrones de filotaxis son 1/2, 1/3, 2/5, 3/8, 5/13, 8/21, etc., donde los numeradores y denominadores forman la secuencia de Fibonacci.
La filotaxis 3/8 se refiere a un patrón de disposición de las hojas en el que cada hoja está separada de la siguiente por tres octavos de una rotación completa de 360 grados alrededor del tallo. Esto significa que cada hoja sucesiva se sitúa en un ángulo de 3/8×360=135 grados (llamado ángulo de divergencia) respecto a la anterior. El ángulo de divergencia converge al ángulo áureo de 137,5 grados en plantas con un gran número de hojas. Esta divergencia fraccional ayuda a distribuir las hojas de forma que se maximice la exposición a la luz solar y se minimicen el solapamiento y la sombra, garantizando que cada hoja reciba la luz y el aire adecuados. Un espaciado adecuado permite una distribución óptima del agua y los nutrientes por toda la planta.

Fig. 3.27. Filotaxis 2/5 (a) y filotaxis 3/8 (b)
También se pueden encontrar patrones similares en muchas flores. Por ejemplo, el número de hojas, ramas y pétalos de la estornudera forma números de Fibonacci consecutivos. 1, 1, 2, 3, 5, 8 para las hojas, 1, 2, 3, 5, 8, 13 para las ramas y 5, 8 u 8, 13 para los pétalos.

Fig. 3.28. Hojas y ramas de estornudaria
No sólo las hojas, sino también los brotes, frutos y semillas de una planta se rigen por la secuencia de Fibonacci y el ángulo áureo.
El patrón de brotación de la pícea sigue los principios de la secuencia de Fibonacci y el ángulo áureo. Cada nuevo brote emerge en un ángulo de aproximadamente 137,5 grados (ángulo áureo) con respecto al anterior.

Fig. 3.29. Patrón de brotación de la pícea
Como resultado, las ramas forman un patrón en espiral alrededor del tronco, alineándose con los números de Fibonacci en su distribución.Este patrón natural mejora la capacidad del árbol para recoger eficazmente la luz solar, el agua y los nutrientes, favoreciendo su crecimiento y salud. Este patrón natural mejora la capacidad del árbol para recoger con eficacia la luz solar, el agua y los nutrientes, favoreciendo su crecimiento y salud.La margarita exhibe el patrón de Fibonacci y el ángulo de oro en su disposición floral. Los pétalos y las semillas de la flor se alinean en espirales que siguen la secuencia de Fibonacci, en la que el número de espirales en cada dirección suele corresponder a números de Fibonacci sucesivos, como el 21 y el 34. Además, el ángulo de divergencia entre los pétalos y las semillas de la margarita es el mismo que el de la margarita. Además, el ángulo de divergencia entre pétalos o semillas sucesivos es aproximadamente el ángulo áureo. Si la espiral se enrolla en un ángulo áureo, forma una espiral logarítmica. Si los floretes de una margarita forman una espiral logarítmica, mantienen su forma a medida que crecen. Una espiral logarítmica es autosimilar, lo que significa que la forma de la espiral se mantiene constante incluso cuando se expande. Las propiedades inherentes a la espiral logarítmica permiten a la margarita mantener su estructura geométrica general durante todo su crecimiento.
Patrones similares se encuentran en las piñas, la coliflor y el brócoli Romanesco. Las escamas de una piña están dispuestas en espirales que siguen los números de Fibonacci, generalmente 8 espirales en una dirección y 13 en la dirección opuesta, con cada escama cuidadosamente colocada aproximadamente en el ángulo áureo. Del mismo modo, los ramilletes de la coliflor se enrollan en 5 espirales en una dirección y 8 en la otra, reflejando la misma secuencia numérica. En el brécol Romanesco, los floretes se disponen en 13 espirales en una dirección y 21 en la otra.
Los números de Fibonacci de las piñas se encuentran en la disposición de sus ojos. Estos ojos están organizados en espirales que siguen los números de Fibonacci, formando normalmente tres conjuntos distintos de espirales. Comúnmente, se pueden encontrar 8 espirales ascendentes en una dirección, 13 en la dirección opuesta, y a veces 21 en otra, cada conjunto alineado con números de Fibonacci consecutivos. Este patrón garantiza un envasado eficaz y maximiza la integridad estructural de la fruta. La disposición permite que la piña crezca uniformemente y distribuya los nutrientes por igual, mostrando la aplicación natural de las secuencias de Fibonacci en el crecimiento y desarrollo de las plantas.

Fig. 3.30. Secuencia de Fibonacci y espiral logarítmica en plantas
La curva de crecimiento que sigue una espiral logarítmica se encuentra no sólo en las plantas, sino también en los seres humanos y otros animales. Algunos ejemplos son el pabellón auricular humano, la cóclea del oído, los dedos humanos, la cola del caballito de mar, los cuernos de la cabra montés y las conchas de varios caracoles, incluido el nautilus. Si estos patrones de crecimiento no siguieran una espiral logarítmica, serían incapaces de mantener su forma característica a medida que siguen creciendo, perdiendo en última instancia su funcionalidad distintiva y su integridad estructural única.
Por ejemplo, si el patrón de crecimiento de la cóclea no siguiera una espiral logarítmica, afectaría significativamente a su capacidad para procesar el sonido de forma eficaz. La espiral logarítmica permite detectar un gradiente de frecuencias a lo largo de su longitud, con frecuencias altas en la base y frecuencias bajas en el ápice. Las desviaciones de este patrón podrían dar lugar a un espaciado desigual de las zonas de detección de frecuencias, lo que provocaría una audición deficiente o dificultades para distinguir entre distintas frecuencias sonoras. Esta disposición precisa es esencial para que la cóclea convierta las ondas sonoras en señales neuronales, lo que permite una percepción auditiva precisa.

Fig. 3.31. Cóclea, oreja, caballito de mar y hueso del nudillo de la mano.
En la naturaleza se pueden encontrar muchos patrones fractales, como los patrones de ramificación de helechos y árboles, la estructura de las hojas de los helechos, la disposición de los ramilletes en la coliflor, el brécol y el brécol romanesco, los sistemas radiculares de muchas plantas y las piñas. Los patrones fractales también están presentes en los sistemas biológicos.
La ramificación de los vasos sanguíneos, desde las grandes arterias hasta los capilares más pequeños, sigue patrones fractales. La estructura fractal maximiza la superficie de intercambio de nutrientes y gases, al tiempo que minimiza la energía necesaria para bombear la sangre por todo el cuerpo. La ramificación fractal garantiza que cada célula reciba suficiente oxígeno y nutrientes. Además, la naturaleza fractal de los vasos sanguíneos contribuye a su robustez y adaptabilidad. Los patrones repetitivos pueden adaptarse fácilmente al crecimiento y la reparación, manteniendo una circulación eficiente a pesar de los cambios o daños.
El sistema respiratorio humano también presenta patrones fractales. La estructura del pulmón comprende la tráquea que se ramifica en bronquios, que a su vez se dividen en bronquiolos más pequeños y culminan en alvéolos donde se produce el intercambio gaseoso. Cada división mantiene patrones fractales. Esta arquitectura fractal maximiza la superficie, que es tan grande como el tamaño de una pista de tenis, para el intercambio de gases, al tiempo que minimiza el volumen ocupado por los pulmones. Al seguir un patrón fractal, los pulmones pueden suministrar eficazmente oxígeno al torrente sanguíneo y expulsar dióxido de carbono, optimizando la función respiratoria.

Fig. 3.32. Fractales encontrados en el helecho y el románico
La presencia de patrones matemáticos como el ángulo de oro, la secuencia de Fibonacci y los fractales en la naturaleza y los sistemas biológicos cuestiona la idea de las mutaciones aleatorias y la selección natural. El espaciado óptimo de las hojas del ángulo de oro y la eficacia de la secuencia de Fibonacci en la disposición de las semillas, por ejemplo, sugieren un diseño intencionado para maximizar la utilización de los recursos. La complejidad autosimilar de los fractales en estructuras como los vasos sanguíneos y las raíces de las plantas indica un sofisticado nivel de organización que no puede lograrse mediante procesos aleatorios. La complejidad, precisión y presencia universal de estas estructuras apuntan a un diseño inteligente predeterminado más que a un proceso evolutivo no dirigido.
4. Invitación a el Evangelio
“Cuando veo tus cielos, obra de tus dedos, La luna y las estrellas que tú formaste,
Digo: ¿Qué es el hombre, para que tengas de él memoria, Y el hijo del hombre, para que lo visites?
Le has hecho poco menor que los ángeles, Y lo coronaste de gloria y de honra.
Le hiciste señorear sobre las obras de tus manos; Todo lo pusiste debajo de sus pies:
Ovejas y bueyes, todo ello, Y asimismo las bestias del campo,
Las aves de los cielos y los peces del mar; Todo cuanto pasa por los senderos del mar.
¡Oh Jehová, Señor nuestro, Cuán grande es tu nombre en toda la tierra!” (Salmo 8:3-9)
Los versículos bíblicos anteriores reflejan bellamente el asombro y la maravilla de la creación, reconociendo la majestuosidad de los cielos y el intrincado diseño del universo como evidencia de un Creador. En estos versículos, el salmista se maravilla ante la luna, las estrellas y la vasta extensión del cielo, que Dios ha colocado en su lugar, reconociendo el acto deliberado e intencionado de la creación. El creacionismo se basa en este sentimiento de asombro, afirmando que la complejidad y el orden que se observan en la naturaleza no son producto del azar, sino del diseño intencionado del Creador divino. La reflexión del salmista sobre la pequeñez de la humanidad en comparación con la grandeza del cosmos pone de relieve la creencia de que, a pesar de la inmensidad del universo, Dios ha elegido coronarnos de gloria y honor, dándonos dominio sobre las obras de Sus manos. Esta profunda relación entre Dios y la humanidad señala a Su profundo amor por nosotros y Su deseo de que vivamos en comunión con Él.
En este capítulo, me gustaría presentar el Evangelio, que revela cómo el amor de Dios y su deseo de comunión con nosotros se cumplen a través de la vida, muerte y resurrección de Jesucristo, ofreciéndonos la oportunidad de reconciliarnos con Él y de vivir en la plenitud de Su gracia. Para aquellos que todavía luchan por creer en la existencia de Dios tal como se revela a través del universo y de toda la creación, también me gustaría presentar la Apuesta de Pascal. Blaise Pascal fue un filósofo, matemático, físico y escritor francés del siglo XVII famoso por sus reflexiones filosóficas sobre la naturaleza humana y la fe, especialmente en su obra ‘Pensées’. Presentó un argumento filosófico sobre la existencia de Dios llamado ‘La apuesta de Pascal’. Pascal sostiene que es una decisión racional vivir como si Dios existiera porque si Dios existe, el creyente obtiene la felicidad eterna, mientras que si Dios no existe, la pérdida es insignificante. Por el contrario, si uno vive como si Dios no existiera y se equivoca, la pérdida potencial es inmensa, incluido el sufrimiento eterno, mientras que la ganancia si acierta es mínima. De ahí que Pascal concluya que creer en Dios es la ‘apuesta’ más segura y beneficiosa.
Creer en Dios | Dios existe | Dios no existe |
Sí | cielo | No pasa nada |
No | infierno | No pasa nada |
Tabla 4.1. La apuesta de Pascal
Hasta ahora, hemos debatido ampliamente sobre la creación y la evolución, reconociendo la existencia de Dios. Si reconoces esta verdad, entonces la Apuesta de Pascal presenta dos opciones claras: la alegría eterna (el cielo) o el sufrimiento eterno (el infierno). Todo el mundo desea elegir la primera opción, y nadie quiere elegir la segunda. A estas alturas, puede que dudes de la existencia del cielo, pero el cielo existe de verdad. En 2 Corintios, el apóstol Pablo comparte una experiencia profunda y misteriosa que permite vislumbrar la existencia del cielo. Escribe:
“Conozco a un hombre en Cristo, que hace catorce años (si en el cuerpo, no lo sé; si fuera del cuerpo, no lo sé; Dios lo sabe) fue arrebatado hasta el tercer cielo. Y conozco al tal hombre (si en el cuerpo, o fuera del cuerpo, no lo sé; Dios lo sabe), que fue arrebatado al paraíso, donde oyó palabras inefables que no le es dado al hombre expresar.” (2 Corintios 12:2-4)
El relato de Pablo sugiere que el cielo, o el ‘tercer cielo’, es un reino de belleza indescriptible y presencia divina, distinto de nuestra experiencia terrenal. Este ‘tercer cielo’ se considera la parte más elevada del cielo, un lugar de máxima realidad espiritual y comunión con Dios. Las ‘cosas inefables’ que Pablo oyó allí indican que las experiencias y verdades del cielo están más allá de la comprensión y el lenguaje humanos.
Este pasaje reafirma a los creyentes en la realidad del cielo y en su naturaleza profunda y trascendente, ofreciendo esperanza y una promesa de los misterios divinos que aguardan más allá de nuestra existencia terrenal. La visión de Pablo sirve de poderoso testimonio de la existencia de un paraíso celestial, un lugar preparado por Dios para quienes le aman.
El Cielo está abierto a cualquiera que crea en Jesucristo. Jesucristo vino a la Tierra para salvar a la humanidad del pecado. Jesús es una figura histórica. Nuestra historia se divide en a.C. (antes de Cristo) y A.D. (Anno Domini, que en latín significa ‘en el año de nuestro SEÑOR’). Como se relata en los cuatro Evangelios, Jesús realizó numerosos milagros durante Su ministerio, demostrando Su poder divino y Su compasión. Curó a los enfermos, como a un leproso (Mateo, 8:1-4) y devolvió la vista a los ciegos (Juan, 9:1-7). También hizo milagros en la naturaleza, como calmar una tormenta (Marcos 4:35-41) y caminar sobre el agua (Mateo 14:22-33). Además, Jesús resucitó a los muertos, sobre todo a Lázaro (Juan 11:1-44), y multiplicó los panes y los peces para alimentar a miles de personas (Mateo 14:13-21). Estos milagros afirmaron Su identidad como Hijo de Dios y trajeron esperanza y fe a muchos.
Si quieres creer en Jesús y buscar la seguridad de ir al cielo, puedes seguir estos pasos basados en los principios fundamentales de la fe cristiana:
Reconoce que eres un pecador que necesita el perdón de Dios. El pecado incluye la blasfemia, el orgullo, la avaricia, la lujuria, la ira, la idolatría, el adulterio, el robo, la mentira, el engaño, el odio, los juegos de azar, la embriaguez y el abuso de drogas, y más-nadie está exento de él. Este pecado ha roto nuestra comunión con Dios, creando una división entre nosotros y Él. La Biblia dice,
“por cuanto todos pecaron, y están destituidos de la gloria de Dios,” (Romanos 3:23).
Ten fe en que Jesucristo es el Hijo de Dios que murió por tus pecados y resucitó.
“Porque de tal manera amó Dios al mundo, que ha dado a su Hijo unigénito, para que todo aquel que en él cree, no se pierda, mas tenga vida eterna.” (Juan 3:16)
Confiesa tus pecados a Dios y apártate de ellos.
“Si confesamos nuestros pecados, él es fiel y justo para perdonar nuestros pecados, y limpiarnos de toda maldad.” (1 Jn 1,9)
Invita a Jesús a tu vida para que sea tu Salvador y Señor. Esto significa confiar en Él para tu salvación y comprometerte a seguirle.
“Mas a todos los que le recibieron, a los que creen en su nombre, les dio potestad de ser hechos hijos de Dios;” (Jn 1,12).
He aquí una sencilla oración que puedes rezar para expresar tu fe y tu compromiso con Jesús:
“Vengo ante Ti, reconociendo mis pecados y mi necesidad de Tu gracia. Creo que Jesús murió por mis pecados y resucitó para darme una nueva vida. Lo acepto como mi Señor y Salvador, rindiéndote mi corazón y mi vida. Por favor, perdóname, límpiame y guíame por Tu Espíritu. Ayúdame a vivir fielmente, caminando en Tu amor y propósito. Gracias por Tu misericordia y salvación. En el nombre de Jesús, Amén.”
Después de aceptar a Jesús, es importante que crezcas en tu nueva fe. Lee la Biblia con regularidad, reza y busca una iglesia local donde puedas formar parte de una comunidad de creyentes que te apoyen y animen.
Demuestra tu fe con tus actos amando a los demás, compartiendo tu fe en y viviendo según las enseñanzas de Jesús.
“En esto conocerán todos que sois mis discípulos, si tuviereis amor los unos con los otros.” (Jn 13,35)
Creer en Jesús y entregarle tu vida es El fundamento de la fe cristiana y el camino hacia la vida eterna en el Cielo.
“Ellos dijeron: Cree en el Señor Jesucristo, y serás salvo, tú y tu casa.” (Hechos 16:31)
Agradecimientos
Quisiera expresar mi sincera gratitud al reverendo Hwan-Chull Park, de la Bridge Church, que leyó detenidamente todo el borrador y realizó meticulosas revisiones y adiciones necesarias.
También estoy profundamente agradecido al reverendo Yong-Cheol Kim, al reverendo Jong-Kug Kim, al misionero Kyoung Kim y a la señora Hyun-Ah Kim por inspirar la publicación de este libro a través de muchas conversaciones sobre la Biblia y la astronomía.
Además, hago extensivo mi más sincero agradecimiento al Dr. y al reverendo Jun-Sub Im, de la iglesia coreana BLOO-gene de Charlottesville, al Dr. Kyoung-Joo Choi, de Arcturus Therapeutics, y al Dr. Chi-Hoon Park, del Instituto Coreano de Investigación de Tecnología Química, por leer el manuscrito y aportar valiosos comentarios
Agradezco especialmente a mis hijos, Samuel y Daniel, su ayuda con el trabajo de imagen.
¡Toda la gloria a Dios!
Crédito de la imagen
1. La creación del Universo
Fig. 1.1: NASA/JPL, Fig. 1.2: Hubble Heritage Team, Fig. 1.3: R. Hurt/JPL-Caltech/NASA, Fig. 1.4: Hubble/NASA/ESA, Fig. 1.5: Wikipedia/R. Powell, Fig. 1.6: Wikimedia/D. Leinweber, Fig. 1.7: NASA/CXC/M. Weiss(izquierda), NASA/D. Berry (derecha), Fig. 1.8: Stellarium, Fig. 1.9: Physics Forums, Fig. 1.10: NASA/JPL-Caltech (izquierda), A. Sarangi, 2018, SSR, 214, 63 (derecha), Fig. 1.11: Wikimedia/ALMA (ESO/NAOJ/NRAO) (izquierda), T. Müller (HdA/MPIA)/G. Perotti (The MINDS collaboration)/M. Benisty (derecha), Fig. 1.12: TASA Graphic Arts, Inc., Fig. 1.14: Jon Therkildsen, Fig. 1.15: www.neot-kedumim.org.il
2. La obra maestra de Dios, la Tierra
Fig. 2.1: R. Narasimha, Fig. 2.3: NASA, Fig. 2.4: NASA/Goddard/Aaron Kaase, Fig. 2.6: Wikimedia, Fig. 2.7: Linda Martel, Fig. 2.8: Wikimedia, Fig. 2.9: NASA/ESA/H. Weaver & E. Smith (izquierda), NASA/HST Comet Team (derecha), Fig. 2.10: Wikimedia/M. Bitton, Fig. 2.11: Wikimedia/John Garrett, Fig. 2.12: UK Foreign and Commonwealth Office, Fig. 2.13: Wikipedia, Fig. 2.16: Wikipedia/G. Taylor, Fig. 2.17: NASA/Caltech
3. ¿Creación o evolución?
Fig. 3.1. Wikipedia/Yassine Mrabet Wikipedia/Yassine Mrabet, Fig. 3.2: OpenEd/Christine Miller, Fig. 3.3: Wikipedia/LadyofHats, Fig. 3.4: Wikipedia/Messer Woland & Szczepan (izquierda), Wikipedia/LadyofHats (derecha), Fig. 3.5: J.E. Duncan & S.B. Goldstein, Fig. 3.6: Wikipedia/Fiona 126, Fig. 3.7: NASA, Fig. 3.8: R. Cui, Fig. 3.9: Wikipedia/Ansgar Walk, Fig. 3.10: The Whisker Chronicles, Fig. 3.11: Encyclopedia Britanica Inc, Fig. 3.12: Wikipedia, Fig. 3.13: Wikipedia/LadyofHats, Fig. 3.14: Wikipedia/J.J. Corneveaux, Fig. 3.15: Smithsonian Institution, Fig. 3.17: NRAO/AUI/NSF (izquierda), Wikipedia/Colby Gutierrez-Kraybill (derecha), Fig. 3.18: Wikipedia/MikeRun, Fig. 3.20 - Fig. 3.23: Shueisha, Inc./Obara Takuya, Fig. 3.24: Wikipedia/Pinakpani, Fig. 3.25: Wikipedia/Dicklyon, Fig. 3.26: Wikipedia/Stannered (1st img), Dicklyon (2nd img), Morn the Gom (3(rd) img), Eequor (4th img), Fig. 3.27: M. Kitazawa/J. Plant Res., Fig. 3.28: S.R. Rahaman, Fig. 3.30: Jill Britton (piña), Fig. 3.32: Wikipedia/Farry (izquierda), Wikimedia/Ivar Leidus (derecha).
Referencias
1. La creación del Universo
제자원 (2002), Oxford Bible Encyclopedia, Bible Textbook Co., Genesis Chap. 1-11.
Another universe existed before Big Bang? 우주먼지의 현자타임즈, 2/24/2024, https://www.youtube.com/watch?v=RckLkaVzFe0
A Big Ring on The Sky: AAS 243rd Press conference. Alexia M. Lopez, 1/11/2024, https://www.youtube.com/watch?v=fwRJGaIcX6A
Bogdan, A., et al. (2024), ‘Evidence for heavy-seed origin of early supermassive black holes from a z ≈ 10 X-ray quasar’, Nature Astronomy, 8, 126.
Bonanno, A., & Fröhlich, H.-E. (2015), ‘A Bayesian estimation of the helioseismic solar age’, Astronomy & Astrophysics, 580, A130.
Karim, M. T., & Mamajek, E. E. (2017), ‘Revised geometric estimates of the North Galactic Pole and the sun's height above the Galactic mid-plane’, MNRAS, 465, 472.
Lopez, A. M., et al. (2022), ‘Giant Arc on the sky’, MNRAS, 516, 1557.
Lopez, A. M., Clowes, R. G., & Williger, G. M. (2024), ‘A Big Ring on the Sky’, JCAP, 07, 55.
Lyra, W., et al. (2023), ‘An Analytical Theory for the Growth from Planetesimals to Planets by Polydisperse Pebble Accretion’, The Astrophysical Journal, 946, 60.
Penrose, R. (2016), The Emperor’s New Mind, Oxford University Press, Oxford, United Kingdom.
Perotti, G., et al. (2023), ‘Water in the terrestrial planet-forming zone of the PDS 70 disk’, Nature, 620, 516.
Sandor, Zs., et al. (2024), ‘Planetesimal and planet formation in transient dust traps’, Astronomy & Astrophysics, in press.
Schiller, M., et al. (2020), ‘Iron isotope evidence for very rapid accretion and differentiation of the proto-earth’, Science Advances, 6, 7.
Tonelli, G. (2019), Genesis: The story of how everything began, Farrah, Straus and Giroux, New York, pp 19-44
Tryon, E. P. (1973), ‘Is the Universe a vacuum fluctuation’, Nature, 246, 396.
Vorobyov, E. I., et al. (2024), ‘Dust growth and pebble formation in the initial stages of protoplanetary disk evolution’, Astronomy & Astrophysics, 683, A202.
Yi, S., et al. (2001), ‘Toward Better Age Estimates for Stellar Populations: The Y2 Isochrones for Solar Mixture’, The Astrophysical Journal Supplement Series, 136, 417.
2. La obra maestra de Dios, la Tierra
Comins, N. F. (1993), What If the Moon Didn't Exist? HarperCollins Publishers Inc., New York, NY.
Gonzalez, G. & Richards, J. W. (2004), The privileged planet: How Our Place in the Cosmos Is Designed for Discovery, Regnery Publishing, Inc.
Lineweaver, C. H., et al. (2004), ‘The Galactic Habitable Zone and the Age Distribution of Complex Life in the Milky Way’, Science, 303 (5654), 59.
Lüthi, D. et al. (2008), ‘High-resolution carbon dioxide concentration record 650,000 - 800,000 years before present’, Nature, 453, 379.
Narasimha, R., et al. (2023), ‘Making Habitable Worlds: Planets Versus Megastructures’, arXiv:2309.06562.
OpenAI. (2024), ChatGPT (4o) [Large language model], https://chatgpt.com
Ward, Peter D. & Brownlee, Donald (2000), Rare Earth: Why Complex Life is Uncommon in the Universe, Copernicus Books (Springer Verlag).
3. ¿Creación o evolución?
Abelson, P. H. (1966), ‘Chemical Events on the Primitive Earth’, Proc Nat Acad Sci, 55, 1365.
Behe, M. J. (2006). Darwin's black box: The biochemical challenge to evolution. Free Press.
Behe, M. J. (2020). Darwin devolves: The new science about DNA that challenges evolution. HarperOne.
Bernhardt, H. S. (2012), ‘The RNA world hypothesis: the worst theory of the early evolution of life (except for all the others)’, Biology Direct, 7, Article number: 23.
Chyba, C. F., & Sagan, C. (1992), ‘Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: An inventory for the origins of life’. Nature, 355, 125.
Cui, R., ‘The transcription network in skin tanning: from p53 to microphthalmia’, https://www.abcam.com/index.html?pageconfig=resource&rid=11180&pid=10026
Dembski, W. A., & Ewert, W. (2023). The design inference: Eliminating chance through small probabilities. Discovery Institute.
Danielson, M. (2020), ‘Simultaneous Determination of L- and D-Amino Acids in Proteins’, Foods, 9 (3), 309.
Fabre, J.-H. (2015), The Mason -Bees (Perfect Library), CreateSpace Independent Publishing Platform.
Higgins, M. (2014), ‘Bear evolution 101’, The Whisker Chronicles, https://thewhiskerchronicles.com/2014/01/03/bear-evolution-101/
Kasting, J. F. (1993). ‘Earth's Early Atmosphere.’ Science, 259(5097), 920.
Maslin, M. (2016), ‘Forty years of linking orbits to ice ages’, Nature, 540 (7632), 208.
Miller, S. L. (1953), ‘A Production of Amino Acids under Possible Primitive Earth Conditions’, Science, 117, 528
Mumma, M. M., et al. (1996), ‘Detection of Abundant Ethane and Methane, Along with Carbon Monoxide and Water, in Comet C/1996 B2 Hyakutake: Evidence for Interstellar Origin’, Science, 272 (5266), 1310.
OpenAI. (2024), ChatGPT (4o) [Large language model], https://chatgpt.com
Park, Chi Hoon (2024), ‘Stop codon points to GOD’, Proceedings of the 20th Anniversary KRAID Symposium
Pinto, J. P., Gladstone, G. R., & Yung, Y. L. (1980), ‘Photochemical Production of Formaldehyde in Earth’s Primitive Atmosphere’, Science, 210, 183.
Pinto, O. H., et al. (2022), ‘A Survey of CO, CO2, and H2O in Comets and Centaurs’, Planet. Sci. J., 3, 247.
Russo, D., et al. (2016), ‘Emerging trends and a comet taxonomy based on the volatile chemistry measured in thirty comets with high resolution infrared spectroscopy between 1997 and 2013’, Icarus, 278, 301.
Sanjuán, R., Moya, A., & Elena, S. F. (2004), ‘The distribution of fitness effects caused by single-nucleotide substitutions in an RNA virus’, Proc Natl Acad Sci, 101(22), 8396.
Trail, D., et al. (2011), ‘The oxidation state of Hadean magmas and implications for early Earth’s atmosphere’, Nature, 480, 79.
Urey, H. C. (1952). ‘On the Early Chemical History of the Earth and the Origin of Life.’ Proc Natl Acad Sci, 38(4), 351.
Wikipedia, Mutation (Distribution of fitness effects).
Wikipedia, Visual phototransduction.
Yang, P.-K. (2016), ‘How does Planck’s constant influence the macroscopic world?’, Eur. J. Phys., 37, 055406.
Zahnle, K. J. (1986), ‘Photochemistry of methane and the formation of hydrocyanic acid (HCN) in the Earth’s early atmosphere’, J. Geophys Res, 91, 2819.
Sobre el autor
El Dr. Dongchan Kim se licenció en Astronomía por la Universidad Yonsei de Seúl (Corea) y se doctoró en Astronomía por la Universidad de Hawai. Tras completar sus estudios de doctorado, realizó investigaciones astronómicas en varias instituciones, entre ellas el Laboratorio de Propulsión a Chorro/Caltech de la NASA, la Universidad Nacional de Seúl y la Universidad de Virginia.
La investigación del Dr. Kim se centra en las galaxias infrarrojas luminosas (LIRG), las galaxias infrarrojas ultraluminosas (ULIRG), los cuásares y los agujeros negros supermasivos en retroceso.
Está afiliado al Observatorio Radioastronómico Nacional de Charlottesville, Virginia, EE.UU.
Este libro es una traducción al español de [DIVINE GENESIS: Exploring Creation through Astronomy and Biology] publicado en inglés por Amazon USA. The PDF version of this book, along with translations in multiple languages, can be downloaded from divine-genesis.org.